Proteinat janë komponime organike me molekulare të lartë. Këto substanca quhen gjithashtu proteina dhe polipeptide. Më pas, le të shohim strukturën dhe funksionet e proteinave.
Informacione të përgjithshme
Struktura kimike e proteinave përfaqësohet nga aminoacide alfa të lidhura në një zinxhir përmes një lidhjeje peptide. Në organizmat e gjallë, përbërja përcaktohet nga kodi gjenetik. Në procesin e sintezës, në shumicën e rasteve përdoren 20 aminoacide të tipit standard. Kombinimet e tyre të shumta formojnë molekula proteinike me një larmi të gjerë vetive. Mbetjet e aminoacideve shpesh i nënshtrohen modifikimeve pas përkthimit. Ato mund të lindin përpara se proteina të fillojë të kryejë funksionet e saj dhe gjatë aktivitetit të saj në qelizë. Në organizmat e gjallë, disa molekula shpesh formojnë komplekse komplekse. Një shembull është lidhja fotosintetike.
Qëllimi i lidhjeve
Proteinat konsiderohen si një komponent i rëndësishëm i ushqimit të njerëzve dhe kafshëve për faktin se trupi i tyre nuk mund të sintetizojë të gjitha aminoacidet e nevojshme. Disa prej tyre duhet të vijnë me ushqime proteinike. Burimet kryesore të komponimeve janë mishi, arrat, qumështi, peshku dhe drithërat. Në një masë më të vogël, proteinat janë të pranishme në perime, kërpudha dhe manaferra. Gjatë tretjes përmes enzimave, proteinat e konsumuara zbërthehen në aminoacide. Ato përdoren tashmë në biosintezën e proteinave të tyre në trup ose i nënshtrohen ndarjes së mëtejshme për të marrë energji.

Sfondi historik
Sekuenca e strukturës së proteinës së insulinës u përcaktua për herë të parë nga Frederij Senger. Për punën e tij ai mori çmimin Nobel në 1958. Sanger përdori metodën e renditjes. Duke përdorur difraksionin me rreze X, strukturat tre-dimensionale të mioglobinës dhe hemoglobinës u morën më pas (në fund të viteve 1950). Puna u krye nga John Kendrew dhe Max Perutz.
Struktura e molekulës së proteinës
Ai përfshin polimere lineare. Ata, nga ana tjetër, përbëhen nga mbetje të aminoacideve alfa, të cilat janë monomere. Përveç kësaj, struktura e proteinave mund të përfshijë përbërës të një natyre jo-aminoacide dhe mbetje të modifikuara të aminoacideve. Gjatë përcaktimit të komponentëve, përdoren shkurtesat me 1 ose 3 shkronja. Një përbërës që përmban nga dy deri në disa dhjetëra mbetje shpesh quhet "polipeptid". Si rezultat i ndërveprimit të grupit alfa-karboksil të një aminoacidi me grupin alfa-amino të një tjetri, shfaqen lidhje (gjatë formimit të strukturës së proteinës). Skajet C- dhe N-terminale të përbërjes dallohen, varësisht se cili grup i mbetjes së aminoacideve është i lirë: -COOH ose -NH2. Në procesin e sintezës së proteinave në ribozom, mbetja e parë përfundimtare është zakonisht një mbetje metionine; ato pasardhëse i shtohen skajit C të paraardhësve.

Nivelet e organizimit
Ata u propozuan nga Lindrem-Lang. Përkundër faktit se kjo ndarje konsiderohet disi e vjetëruar, ajo ende përdoret. U propozua të dallohen katër nivele të organizimit të lidhjes. Struktura primare e një molekule proteine përcaktohet nga kodi gjenetik dhe karakteristikat e gjenit. Nivelet më të larta karakterizohen nga formimi gjatë palosjes së proteinave. Struktura hapësinore e një proteine përcaktohet në tërësi nga zinxhiri i aminoacideve. Sidoqoftë, ai është mjaft i paqëndrueshëm. Mund të ndikohet nga faktorë të jashtëm. Në këtë drejtim, është më e saktë të flasim për konformimin e përbërjes që është më i favorshmi dhe energjikisht më i preferuari.
Niveli 1
Ai përfaqësohet nga një sekuencë e mbetjeve aminoacide të një zinxhiri polipeptid. Si rregull, përshkruhet duke përdorur shënime me një ose tre shkronja. Struktura primare e proteinave karakterizohet nga kombinime të qëndrueshme të mbetjeve të aminoacideve. Ata kryejnë detyra specifike. Të tilla "motive konservatore" mbeten të ruajtura gjatë evolucionit të specieve. Ato shpesh mund të përdoren për të parashikuar problemin e një proteine të panjohur. Duke vlerësuar shkallën e ngjashmërisë (homologjisë) në zinxhirët e aminoacideve nga organizma të ndryshëm, është e mundur të përcaktohet distanca evolucionare e formuar midis taksave që përbëjnë këta organizma. Struktura primare e proteinave përcaktohet nga sekuenca ose nga kompleksi origjinal i mARN-së së saj duke përdorur një tabelë kodi gjenetik.

Renditja lokale e një seksioni zinxhir
Ky është niveli tjetër i organizimit - struktura dytësore e proteinave. Ka disa lloje të tij. Renditja lokale e një pjese të një zinxhiri polipeptid stabilizohet nga lidhjet hidrogjenore. Llojet më të njohura janë:

Struktura hapësinore
Struktura terciare e proteinave përfshin elementë të nivelit të mëparshëm. Ato stabilizohen nga lloje të ndryshme ndërveprimesh. Lidhjet hidrofobike janë të një rëndësie të madhe. Stabilizimi përfshin:
- Ndërveprimet kovalente.
- Lidhjet jonike të formuara midis grupeve anësore të aminoacideve që kanë ngarkesa të kundërta.
- Ndërveprimet e hidrogjenit.
- Lidhjet hidrofobike. Në procesin e bashkëveprimit me elementët përreth H 2 O, proteina paloset në mënyrë që grupet anësore jopolare të aminoacideve të izolohen nga tretësira ujore. Në sipërfaqen e molekulës shfaqen grupe hidrofile (polare).
Struktura terciare e proteinave përcaktohet me metoda të rezonancës magnetike (bërthamore), lloje të caktuara të mikroskopisë dhe metoda të tjera.
Parimi i shtrimit
Hulumtimet kanë treguar se është e përshtatshme të identifikohet një nivel më shumë midis niveleve 2 dhe 3. Quhet "arkitekturë", "motiv shtrimi". Përcaktohet nga pozicioni relativ i përbërësve të strukturës dytësore (fijet beta dhe helikat alfa) brenda kufijve të një globuli kompakt - domeni i proteinave. Mund të ekzistojë në mënyrë të pavarur ose të përfshihet në një proteinë më të madhe së bashku me të tjera të ngjashme. Është vërtetuar se motivet e stilimit janë mjaft konservatore. Ato gjenden në proteina që nuk kanë as marrëdhënie evolucionare dhe as funksionale. Përkufizimi i arkitekturës është baza e klasifikimit racional (fizik).

Organizimi i domenit
Me rregullimin e ndërsjellë të disa zinxhirëve polipeptidë brenda një kompleksi proteinik, formohet struktura kuaternare e proteinave. Elementet që e përbëjnë atë formohen veçmas në ribozome. Vetëm pas përfundimit të sintezës fillon të formohet kjo strukturë proteinike. Mund të përmbajë zinxhirë polipeptidikë të ndryshëm dhe identikë. Struktura kuaternare e proteinave stabilizohet për shkak të të njëjtave ndërveprime si në nivelin e mëparshëm. Disa komplekse mund të përfshijnë disa dhjetëra proteina.
Struktura e proteinave: detyra mbrojtëse
Polipeptidet e citoskeletit, duke vepruar në një farë mënyre si përforcim, i japin shumë organeleve formën e tyre dhe marrin pjesë në ndryshimin e tij. Proteinat strukturore sigurojnë mbrojtje për trupin. Për shembull, kolagjeni është një proteinë e tillë. Ajo formon bazën në substancën ndërqelizore të indeve lidhëse. Keratina gjithashtu ka një funksion mbrojtës. Ajo formon bazën e brirëve, puplave, flokëve dhe derivateve të tjera të epidermës. Kur proteinat lidhin toksinat, në shumë raste ndodh detoksifikimi. Kështu realizohet detyra e mbrojtjes kimike të organizmit. Enzimat e mëlçisë luajnë një rol veçanërisht të rëndësishëm në procesin e neutralizimit të toksinave në trupin e njeriut. Ata janë në gjendje të zbërthejnë helmet ose t'i kthejnë ato në formë të tretshme. Kjo lehtëson transportin më të shpejtë të tyre nga trupi. Proteinat e pranishme në gjak dhe lëngje të tjera të trupit sigurojnë mbrojtje imune, duke shkaktuar një përgjigje si ndaj sulmit patogjen ashtu edhe ndaj dëmtimit. Imunoglobulinat (antitrupat dhe përbërësit e sistemit të komplementit) janë në gjendje të neutralizojnë bakteret, proteinat e huaja dhe viruset.
Mekanizmi rregullator
Molekulat e proteinave, të cilat nuk veprojnë as si burim energjie dhe as si material ndërtimi, kontrollojnë shumë procese ndërqelizore. Kështu, falë tyre, rregullohet përkthimi, transkriptimi, prerja dhe aktiviteti i polipeptideve të tjera. Mekanizmi rregullues bazohet në aktivitetin enzimatik ose manifestohet për shkak të lidhjes specifike me molekulat e tjera. Për shembull, faktorët e transkriptimit, polipeptidet aktivizues dhe proteinat represore janë në gjendje të kontrollojnë intensitetin e transkriptimit të gjeneve. Duke vepruar kështu, ato ndërveprojnë me sekuencat rregullatore të gjeneve. Roli më i rëndësishëm në kontrollin e rrjedhës së proceseve ndërqelizore u caktohet fosfatazave proteinike dhe kinazave proteinike. Këto enzima nxisin ose pengojnë aktivitetin e proteinave të tjera duke shtuar ose hequr grupe fosfate prej tyre.

Detyrë sinjalizuese
Shpesh kombinohet me funksionin rregullator. Kjo për faktin se shumë polipeptide ndërqelizore, si dhe jashtëqelizore, mund të transmetojnë sinjale. Faktorët e rritjes, citokinat, hormonet dhe komponimet e tjera e kanë këtë aftësi. Steroidet transportohen përmes gjakut. Ndërveprimi i hormonit me receptorin vepron si një sinjal që shkakton përgjigjen e qelizave. Steroidet kontrollojnë përmbajtjen e komponimeve në gjak dhe qeliza, riprodhimin, rritjen dhe proceset e tjera. Një shembull është insulina. Rregullon nivelet e glukozës. Ndërveprimi i qelizave kryhet përmes komponimeve të proteinave sinjalizuese të transmetuara përmes substancës ndërqelizore.
Transporti i elementeve
Proteinat e tretshme të përfshira në lëvizjen e molekulave të vogla kanë një afinitet të lartë për substratin, i cili është i pranishëm në përqendrim të shtuar. Ata gjithashtu kanë aftësinë ta lëshojnë lehtësisht në zonat ku përmbajtja e tij është e ulët. Një shembull është hemoglobina e proteinës së transportit. Ai lëviz oksigjenin nga mushkëritë në inde të tjera dhe prej tyre transferon dioksid karboni. Disa proteina të membranës përfshihen gjithashtu në transportin e molekulave të vogla përmes mureve qelizore, duke i ndryshuar ato. Shtresa lipidike e citoplazmës është e papërshkueshme nga uji. Kjo parandalon përhapjen e molekulave të ngarkuara ose polare. Lidhjet e transportit të membranës zakonisht ndahen në bartës dhe kanale.
Lidhje rezervë
Këto proteina formojnë të ashtuquajturat rezerva. Ato grumbullohen, për shembull, në farat e bimëve dhe vezët e kafshëve. Proteinat e tilla veprojnë si një burim rezervë i materies dhe energjisë. Disa komponime përdoren nga trupi si një rezervuar aminoacidesh. Ata, nga ana tjetër, janë pararendës të substancave aktive të përfshira në rregullimin e metabolizmit.
Receptorët qelizorë
Proteinat e tilla mund të vendosen ose drejtpërdrejt në citoplazmë ose të ngulitura në mur. Një pjesë e lidhjes merr sinjalin. Si rregull, është një substancë kimike, dhe në disa raste një efekt mekanik (shtrirje, për shembull), dritë dhe stimuj të tjerë. Në procesin e ekspozimit të një sinjali ndaj një fragmenti të caktuar të molekulës - receptorit polipeptid - fillojnë ndryshimet e tij konformative. Ato provokojnë një ndryshim në konformacionin e pjesës tjetër që transmeton stimulin tek komponentët e tjerë të qelizës. Dërgimi i një sinjali mund të bëhet në mënyra të ndryshme. Disa receptorë janë të aftë të katalizojnë një reaksion kimik, ndërsa të tjerët veprojnë si kanale jonike që mbyllen ose hapen nën ndikimin e një stimuli. Disa komponime lidhin në mënyrë specifike molekulat e dërguara brenda qelizës.
Polipeptidet motorike
Ekziston një klasë e tërë proteinash që sigurojnë lëvizjen e trupit. Proteinat motorike janë të përfshira në tkurrjen e muskujve, lëvizjen e qelizave dhe aktivitetin e flagjelave dhe qerpikëve. Ato gjithashtu ofrojnë transport të drejtuar dhe aktiv. Kinesinat dhe dyneinat transportojnë molekulat përgjatë mikrotubulave duke përdorur hidrolizën ATP si burim energjie. Këto të fundit lëvizin organelet dhe elementët e tjerë drejt centrozomit nga zonat periferike qelizore. Kinesinat lëvizin në drejtim të kundërt. Dyneinet janë gjithashtu përgjegjëse për aktivitetin e flagjelave dhe qerpikëve.
Struktura hapësinore nuk varet nga gjatësia e zinxhirit polipeptid, por nga sekuenca e mbetjeve të aminoacideve specifike për secilën proteinë, si dhe nga radikalet anësore karakteristike të aminoacideve përkatëse.Struktura hapësinore tre-dimensionale ose konformimi i makromolekulave të proteinave formohet kryesisht nga lidhjet hidrogjenore, si dhe nga ndërveprimet hidrofobike ndërmjet radikaleve anësore jopolare të aminoacideve. Lidhjet e hidrogjenit luajnë një rol të madh në formimin dhe mirëmbajtjen e strukturës hapësinore të makromolekulës së proteinës. Sa i përket ndërveprimeve hidrofobike, ato lindin si rezultat i kontaktit midis radikalëve jopolare që nuk janë në gjendje të thyejnë lidhjet hidrogjenore midis molekulave të ujit, të cilat zhvendosen në sipërfaqen e globulës së proteinës. Ndërsa sinteza e proteinave vazhdon, grupet kimike jopolare grumbullohen brenda globulës dhe ato polare detyrohen të dalin në sipërfaqen e saj. Kështu, një molekulë proteine mund të jetë neutrale, e ngarkuar pozitivisht ose negativisht, në varësi të pH të tretësit dhe grupeve jonogjene në proteinë. Ndërveprimet e dobëta përfshijnë gjithashtu lidhjet jonike dhe ndërveprimet van der Waals. Përveç kësaj, konformimi i proteinave ruhet nga lidhjet kovalente S-S , i formuar midis dy mbetjeve të cisteinës. Si rezultat i ndërveprimeve hidrofobike dhe hidrofile, molekula e proteinës merr spontanisht një ose më shumë nga konformacionet më të favorshme termodinamikisht dhe nëse konformimi vendas prishet si rezultat i ndonjë ndikimi të jashtëm, është i mundur restaurimi i plotë ose pothuajse i plotë i saj.Kështu, konformimi i proteinave është një strukturë tre-dimensionale dhe si rezultat i formimit të saj, shumë atome të vendosura në pjesë të largëta të zinxhirit polipeptid afrohen dhe, duke ndikuar njëri-tjetrin, fitojnë veti të reja që mungojnë në aminoacide individuale ose polipeptide të vogla. Ky është i ashtuquajturi struktura terciare, karakterizohet nga orientimi i vargjeve polipeptidike në hapësirë. Struktura terciare e proteinave globulare dhe fibrilare ndryshon ndjeshëm nga njëra-tjetra. Është zakon të karakterizohet forma e një molekule proteine nga një tregues i tillë si shkalla e asimetrisë (raporti i boshtit të gjatë të molekulës me atë të shkurtër). Për proteinat globulare, shkalla e asimetrisë është 3-5 për proteinat fibrilare, kjo vlerë është shumë më e lartë (nga 80 në 150). Hipoteza e rruzullit të shkrirë. Brenda këtij koncepti, dallohen disa faza të vetë-montimit të proteinave.1. Në një zinxhir polipeptid të shpalosur, me ndihmën e lidhjeve hidrogjenore dhe ndërveprimeve hidrofobike, formohen seksione individuale të strukturës dytësore, të cilat shërbejnë si fara për formimin e strukturave të plota dytësore dhe supersekondare.2. Kur numri i këtyre seksioneve arrin një vlerë të caktuar pragu, ndodh një riorientim i radikaleve anësore dhe zinxhiri polipeptid kalon në një formë të re, më kompakte dhe numri i lidhjeve jokovalente rritet ndjeshëm. Një tipar karakteristik i kësaj faze është formimi i kontakteve specifike midis atomeve të vendosura në pjesë të largëta të zinxhirit polipeptid, por të afruara si rezultat i formimit të strukturës terciare.3. Në fazën e fundit formohet konformacioni amtar i molekulës së proteinës, i shoqëruar me mbylljen e lidhjeve disulfide dhe stabilizimin përfundimtar të konformacionit proteinik. Enzima e dytë që katalizon formimin dhe izomerizimin e lidhjeve disulfide është Izomeraza disulfide e proteinave, funksioni i të cilit përfshin edhe ndarjen e urave disulfide të formuara gabimisht.Përveç kësaj, qelizat përmbajnë një numër proteinash katalitike joaktive, të cilat megjithatë japin një kontribut të madh në formimin e strukturave të proteinave hapësinore. Këto janë të ashtuquajturat chapirones dhe chapironinsKapironet ndihmojnë në montimin e saktë të konformacionit tredimensional të proteinave duke formuar komplekse të kthyeshme jokovalente me një zinxhir polipeptid pjesërisht të palosur, ndërsa njëkohësisht pengojnë lidhjet e formuara gabimisht që çojnë në formimin e strukturave të proteinave funksionalisht joaktive. Lista e funksioneve karakteristike të kapironeve përfshin mbrojtjen e globulave të shkrirë nga grumbullimi, si dhe transferimin e proteinave të saposintetizuara në lokacione të ndryshme qelizore. Kapironet janë kryesisht proteina të goditjes nga nxehtësia, sinteza e të cilave rritet ndjeshëm nën ndikimet stresuese të temperaturës, prandaj quhen edhe hsp (proteina të goditjes nga nxehtësia ). Familjet e këtyre proteinave gjenden në qelizat mikrobike, bimore dhe shtazore. Klasifikimi i kapironëve bazohet në peshën e tyre molekulare, e cila varion nga 10 në 90 kDa . Në thelb, funksionet e kapironeve dhe kapironinave ndryshojnë, megjithëse të dyja janë proteina që ndihmojnë në formimin e strukturës tredimensionale të proteinave. Kapironët e mbajnë zinxhirin polipeptid të saposintetizuar në një gjendje të shpalosur, duke e penguar atë të paloset në një formë të ndryshme nga ajo vendase, dhe kapironinat sigurojnë kushtet për të vetmen strukturë proteinike të saktë, vendase.Ekzistojnë katër nivele të organizimit strukturor të proteinave: primar, sekondar, terciar dhe kuaternar. Çdo nivel ka karakteristikat e veta.
Struktura primare e proteinave është një zinxhir polipeptid linear i aminoacideve të lidhur me lidhje peptide. Struktura primare është niveli më i thjeshtë i organizimit strukturor të një molekule proteine. Stabilitet i lartë i jepet nga lidhjet peptide kovalente midis grupit α-amino të një aminoacidi dhe grupit α-karboksil të një aminoacidi tjetër. [shfaq] .
Nëse grupi imino i prolinës ose hidroksiprolinës është i përfshirë në formimin e një lidhjeje peptide, atëherë ai ka një formë tjetër. [shfaq] .

Kur formohen lidhjet peptide në qeliza, fillimisht aktivizohet grupi karboksil i një aminoacidi dhe më pas bashkohet me grupin amino të një tjetri. Sinteza laboratorike e polipeptideve kryhet afërsisht në të njëjtën mënyrë.
Një lidhje peptide është një fragment i përsëritur i një zinxhiri polipeptid. Ka një numër karakteristikash që ndikojnë jo vetëm në formën e strukturës parësore, por edhe në nivelet më të larta të organizimit të zinxhirit polipeptid:


- bashkëplanariteti - të gjithë atomet e përfshirë në grupin peptid janë në të njëjtin plan;
- aftësia për të ekzistuar në dy forma rezonance (forma keto ose enol);
- pozicioni trans i zëvendësuesve në lidhje me lidhjen C-N;
- aftësia për të formuar lidhje hidrogjeni, dhe secili nga grupet peptide mund të formojë dy lidhje hidrogjenore me grupe të tjera, duke përfshirë ato peptide.
Përjashtim bëjnë grupet peptide që përfshijnë grupin amino të prolinës ose hidroksiprolinës. Ata janë në gjendje të formojnë vetëm një lidhje hidrogjeni (shih më lart). Kjo ndikon në formimin e strukturës dytësore të proteinës. Zinxhiri polipeptid në zonën ku ndodhet prolina ose hidroksiprolina përkulet lehtësisht, pasi nuk mbahet, si zakonisht, nga një lidhje e dytë hidrogjeni.
Nomenklatura e peptideve dhe polipeptideve . Emri i peptideve përbëhet nga emrat e aminoacideve përbërëse të tyre. Dy aminoacide bëjnë një dipeptid, tre bëjnë një tripeptid, katër bëjnë një tetrapeptid, etj. Çdo peptid ose zinxhir polipeptid i çdo gjatësie ka një aminoacid N-terminal që përmban një amino grup të lirë dhe një aminoacid C-terminal që përmban një karboksil të lirë grup. Kur emërtohen polipeptidet, të gjitha aminoacidet renditen në mënyrë sekuenciale, duke filluar me atë N-terminal, duke zëvendësuar në emrat e tyre, përveç atij C-terminal, prapashtesën -in me -yl (pasi aminoacidet në peptide nuk kanë më një grup karboksil, por një karbonil). Për shembull, emri i treguar në Fig. 1 tripeptid - leuc llum fenilalani llum threon në.
Karakteristikat e strukturës primare të proteinës . Në shtyllën kurrizore të zinxhirit polipeptid, strukturat e ngurtë (grupe peptide të sheshta) alternohen me rajone relativisht të lëvizshme (-CHR), të cilat janë të afta të rrotullohen rreth lidhjeve. Tipare të tilla strukturore të zinxhirit polipeptid ndikojnë në rregullimin e tij hapësinor.
Struktura dytësore është një mënyrë e palosjes së një zinxhiri polipeptid në një strukturë të renditur për shkak të formimit të lidhjeve hidrogjenore midis grupeve peptide të të njëjtit zinxhir ose zinxhirëve polipeptidë ngjitur. Sipas konfigurimit të tyre, strukturat dytësore ndahen në spirale (α-spiralja) dhe me shtresa-palosje (strukturë β dhe formë tërthore β).


α-Heliks. Ky është një lloj strukture proteine dytësore që duket si një spirale e rregullt, e formuar për shkak të lidhjeve hidrogjenore ndërpeptide brenda një zinxhiri polipeptid. Modeli i strukturës së α-spiralës (Fig. 2), i cili merr parasysh të gjitha vetitë e lidhjes peptide, u propozua nga Pauling dhe Corey. Karakteristikat kryesore të α-spiralës:
- konfigurimi spirale i zinxhirit polipeptid që ka simetri spirale;
- formimi i lidhjeve hidrogjenore midis grupeve peptide të secilës mbetje të parë dhe të katërt të aminoacideve;
- rregullsia e kthesave spirale;
- ekuivalenca e të gjitha mbetjeve të aminoacideve në α-spiralën, pavarësisht nga struktura e radikaleve të tyre anësore;
- radikalet anësore të aminoacideve nuk marrin pjesë në formimin e heliksit α.
Nga jashtë, a-spiralja duket si një spirale pak e shtrirë e një sobë elektrike. Rregullsia e lidhjeve hidrogjenore midis grupit të parë dhe të katërt peptid përcakton rregullsinë e kthesave të zinxhirit polipeptid. Lartësia e një kthese, ose hapi i spirales α, është 0,54 nm; ai përfshin 3,6 mbetje aminoacide, d.m.th., secila mbetje aminoacide lëviz përgjatë boshtit (lartësia e një mbetjeje aminoacidi) me 0,15 nm (0,54:3,6 = 0,15 nm), gjë që na lejon të flasim për ekuivalencën e të gjitha mbetjeve të aminoacideve në α-spiralën. Periudha e rregullsisë së një α-helix është 5 kthesa ose 18 mbetje aminoacide; gjatësia e një periode është 2.7 nm. Oriz. 3. Modeli a-helix Pauling-Corey
β-Struktura. Ky është një lloj strukture dytësore që ka një konfigurim pak të lakuar të zinxhirit polipeptid dhe formohet nga lidhje hidrogjenore ndërpeptide brenda seksioneve individuale të një zinxhiri polipeptid ose zinxhirëve polipeptidë ngjitur. Quhet gjithashtu një strukturë e palosshme me shtresa. Ka lloje të strukturave β. Rajonet me shtresa të kufizuara të formuara nga një zinxhir polipeptid i një proteine quhen forma ndër-β (strukturë e shkurtër β). Lidhjet hidrogjenore në formën kryq-β formohen midis grupeve peptide të sytheve të zinxhirit polipeptid. Një lloj tjetër - struktura e plotë β - është karakteristikë për të gjithë zinxhirin polipeptid, i cili ka një formë të zgjatur dhe mbahet nga lidhje hidrogjenore ndërpeptide midis zinxhirëve polipeptidikë paralelë ngjitur (Fig. 3). Kjo strukturë i ngjan shakullit të një fizarmonikëje. Për më tepër, variantet e strukturave β janë të mundshme: ato mund të formohen nga zinxhirë paralelë (skajet N-terminale të zinxhirëve polipeptidë drejtohen në të njëjtin drejtim) dhe antiparalele (skajet e terminalit N drejtohen në drejtime të ndryshme). Radikalet anësore të një shtrese vendosen ndërmjet radikaleve anësore të një shtrese tjetër.
Në proteinat, kalimet nga struktura α në b-strukturat dhe mbrapa janë të mundshme për shkak të rirregullimit të lidhjeve hidrogjenore. Në vend të lidhjeve të rregullta hidrogjenore ndërpeptide përgjatë zinxhirit (në sajë të së cilës zinxhiri polipeptid është i përdredhur në një spirale), seksionet spirale hapen dhe lidhjet hidrogjenore mbyllen midis fragmenteve të zgjatura të zinxhirëve polipeptidikë. Ky tranzicion gjendet në keratin, proteina e flokëve. Kur lani flokët me detergjentë alkaline, struktura spirale e β-keratinës shkatërrohet lehtësisht dhe ajo shndërrohet në α-keratinë (flokët kaçurrelë drejtohen).
Shkatërrimi i strukturave të rregullta dytësore të proteinave (a-helika dhe β-struktura), në analogji me shkrirjen e një kristali, quhet "shkrirja" e polipeptideve. Në këtë rast, lidhjet e hidrogjenit prishen dhe zinxhirët polipeptidë marrin formën e një lëmshi të rastësishëm. Rrjedhimisht, qëndrueshmëria e strukturave dytësore përcaktohet nga lidhjet hidrogjenore interpeptide. Llojet e tjera të lidhjeve pothuajse nuk marrin pjesë në këtë, me përjashtim të lidhjeve disulfide përgjatë zinxhirit polipeptid në vendndodhjet e mbetjeve të cisteinës. Peptidet e shkurtra mbyllen në cikle për shkak të lidhjeve disulfide. Shumë proteina përmbajnë si rajone α-spiralë ashtu edhe struktura β. Pothuajse nuk ka proteina natyrale që përbëhen nga 100% spirale α (përjashtim bën paramiozina, një proteinë muskulore që është 96-100% α-spira), ndërsa polipeptidet sintetike kanë 100% spirale.
Proteinat e tjera kanë shkallë të ndryshme të mbështjelljes. Një frekuencë e lartë e strukturave α-spiralore vërehet në paramiozinë, mioglobinë dhe hemoglobinë. Në të kundërt, në tripsinë, një ribonukleazë, një pjesë e rëndësishme e zinxhirit polipeptid paloset në struktura β-shtresore. Proteinat e indeve mbështetëse: keratina (proteina e flokëve, leshi), kolagjeni (proteina e tendinave, e lëkurës), fibroina (proteina e mëndafshit natyral) kanë një konfigurim β të zinxhirëve polipeptidikë. Shkallët e ndryshme të helicitetit të zinxhirëve polipeptidikë të proteinave tregojnë se, padyshim, ka forca që prishin pjesërisht helicitetin ose "thyejnë" palosjen e rregullt të zinxhirit polipeptid. Arsyeja për këtë është një palosje më kompakte e zinxhirit polipeptid proteinik në një vëllim të caktuar, d.m.th., në një strukturë terciare.
Struktura terciare e proteinave
Struktura terciare e një proteine është mënyra se si zinxhiri polipeptid është rregulluar në hapësirë. Në bazë të formës së strukturës së tyre terciare, proteinat ndahen kryesisht në globulare dhe fibrilare. Proteinat globulare më së shpeshti kanë një formë elipsoidale, dhe proteinat fibrilare (si fije) kanë një formë të zgjatur (formë shufre ose bosht).
Sidoqoftë, konfigurimi i strukturës terciare të proteinave nuk jep ende arsye për të menduar se proteinat fibrilare kanë vetëm një strukturë β, dhe proteinat globulare kanë një strukturë α-spiral. Ka proteina fibrilare që kanë një strukturë dytësore të palosur spirale dhe jo me shtresa. Për shembull, α-keratina dhe paramiozina (proteina e muskulit obturator të molusqeve), tropomyosin (proteinat e muskujve skeletorë) i përkasin proteinave fibrilare (kanë formë shufre) dhe struktura e tyre dytësore është α-spiralja; në të kundërt, proteinat globulare mund të përmbajnë një numër të madh të strukturave β.
Spiralizimi i një zinxhiri polipeptid linear zvogëlon madhësinë e tij përafërsisht 4 herë; dhe paketimi në strukturën terciare e bën atë dhjetëra herë më kompakt se zinxhiri origjinal.
Lidhje që stabilizojnë strukturën terciare të një proteine . Lidhjet ndërmjet radikaleve anësore të aminoacideve luajnë një rol në stabilizimin e strukturës terciare. Këto lidhje mund të ndahen në:
- i fortë (kovalent) [shfaq]
.
Lidhjet kovalente përfshijnë lidhjet disulfide (-S-S-) ndërmjet radikaleve anësore të cisteineve të vendosura në pjesë të ndryshme të zinxhirit polipeptid; izopeptid, ose pseudopeptid, - midis grupeve amino të radikaleve anësore të lizinës, argininës dhe jo grupeve α-amino, dhe grupeve COOH të radikaleve anësore të acideve aspartike, glutamike dhe aminocitrike, dhe jo grupeve α-karboksil të aminoacideve. Prandaj emri i këtij lloji të lidhjes - i ngjashëm me peptidin. Një lidhje e rrallë esterike formohet nga grupi COOH i aminoacideve dikarboksilike (aspartik, glutamik) dhe grupi OH i hidroksiaminoacideve (serina, treonina).
- i dobët (polar dhe van der Waals) [shfaq]
.
TE lidhjet polare përfshijnë hidrogjenin dhe jonik. Lidhjet hidrogjenore, si zakonisht, ndodhin midis grupit -NH 2, -OH ose -SH të radikalit anësor të një aminoacidi dhe grupit karboksil të një tjetri. Lidhjet jonike ose elektrostatike formohen kur grupet e ngarkuara të radikalëve anësor -NH + 3 (lizina, arginina, histidina) dhe -COO - (acidet aspartike dhe glutamike) vijnë në kontakt.
Lidhje jo polare, ose van der Waals formohet midis radikaleve hidrokarbure të aminoacideve. Radikalët hidrofobikë të aminoacideve alaninë, valinë, izoleucinë, metioninë, fenilalaninë ndërveprojnë me njëri-tjetrin në një mjedis ujor. Lidhjet e dobëta van der Waals nxisin formimin e një bërthame hidrofobike të radikaleve jopolare brenda globulës së proteinave. Sa më shumë aminoacide jopolare të ketë, aq më i madh është roli i lidhjeve van der Waals në palosjen e zinxhirit polipeptid.
Lidhjet e shumta ndërmjet radikaleve anësore të aminoacideve përcaktojnë konfigurimin hapësinor të molekulës së proteinës.
Karakteristikat e organizimit të strukturës terciare të proteinave . Konformimi i strukturës terciare të zinxhirit polipeptid përcaktohet nga vetitë e radikaleve anësore të aminoacideve të përfshira në të (të cilat nuk kanë një efekt të dukshëm në formimin e strukturave parësore dhe dytësore) dhe mikromjedisit, d.m.th. mjedisi. Kur paloset, zinxhiri polipeptid i një proteine tenton të marrë një formë energjike të favorshme, e karakterizuar nga një minimum energjie të lirë. Prandaj, grupet R jopolare, duke "shmangur" ujin, formojnë, si të thuash, pjesën e brendshme të strukturës terciare të proteinës, ku ndodhet pjesa kryesore e mbetjeve hidrofobike të zinxhirit polipeptid. Nuk ka pothuajse asnjë molekula uji në qendër të globulës së proteinave. Grupet R polare (hidrofile) të aminoacidit ndodhen jashtë kësaj bërthame hidrofobike dhe janë të rrethuar nga molekula uji. Zinxhiri polipeptid është i përkulur në mënyrë të ndërlikuar në hapësirën tre-dimensionale. Kur përkulet, konformimi spirale dytësor prishet. Zinxhiri "prihet" në pikat e dobëta ku ndodhen prolina ose hidroksiprolina, pasi këto aminoacide janë më të lëvizshme në zinxhir, duke formuar vetëm një lidhje hidrogjeni me grupet e tjera peptide. Një vend tjetër i përkuljes është glicina, e cila ka një grup të vogël R (hidrogjen). Prandaj, grupet R të aminoacideve të tjera, kur grumbullohen, priren të zënë hapësirën e lirë në vendndodhjen e glicinës. Një numër aminoacidesh - alanina, leucina, glutamati, histidina - kontribuojnë në ruajtjen e strukturave të qëndrueshme spirale në proteina, dhe si metionina, valina, izoleucina, acidi aspartik favorizojnë formimin e strukturave β. Në një molekulë proteine me një konfigurim terciar, ka rajone në formën e α-spiraleve (spiralja), strukturave β (shtresore) dhe një spirale të rastësishme. Vetëm rregullimi i saktë hapësinor i proteinës e bën atë aktive; shkelja e tij çon në ndryshime në vetitë e proteinave dhe humbje të aktivitetit biologjik.
Struktura e proteinave kuaternare
Proteinat e përbëra nga një zinxhir polipeptid kanë vetëm strukturë terciare. Këto përfshijnë mioglobinën - një proteinë e indit muskulor të përfshirë në lidhjen e oksigjenit, një sërë enzimash (lizozima, pepsina, tripsina, etj.). Megjithatë, disa proteina ndërtohen nga disa zinxhirë polipeptidikë, secila prej të cilave ka një strukturë terciare. Për proteina të tilla është futur koncepti i strukturës kuaternare, i cili është organizimi i disa zinxhirëve polipeptidikë me strukturë terciare në një molekulë të vetme funksionale proteinike. Një proteinë e tillë me strukturë kuaternare quhet oligomer dhe vargjet e saj polipeptide me strukturë terciare quhen protomerë ose nënnjësi (Fig. 4).

Në nivelin kuaternar të organizimit, proteinat ruajnë konfigurimin bazë të strukturës terciare (globulare ose fibrilare). Për shembull, hemoglobina është një proteinë me strukturë kuaternare dhe përbëhet nga katër nënnjësi. Secila prej nënnjësive është një proteinë globulare dhe, në përgjithësi, hemoglobina ka gjithashtu një konfigurim globular. Proteinat e flokëve dhe të leshit - keratinat, të lidhura në strukturën terciare me proteinat fibrilare, kanë një konformacion fibrilar dhe një strukturë kuaternare.
Stabilizimi i strukturës kuaternare të proteinave . Të gjitha proteinat që kanë një strukturë kuaternare janë të izoluara në formën e makromolekulave individuale që nuk ndahen në nënnjësi. Kontaktet ndërmjet sipërfaqeve të nënnjësive janë të mundshme vetëm për shkak të grupeve polare të mbetjeve të aminoacideve, pasi gjatë formimit të strukturës terciare të secilit prej zinxhirëve polipeptidikë, radikalet anësore të aminoacideve jopolare (të cilat përbëjnë shumicën e të gjitha aminoacidet proteinogjene) janë të fshehura brenda nënnjësisë. Midis grupeve të tyre polare formohen lidhje të shumta jonike (kripë), hidrogjeni dhe në disa raste disulfide, të cilat i mbajnë fort nënnjësitë në formën e një kompleksi të organizuar. Përdorimi i substancave që thyejnë lidhjet hidrogjenore ose substancave që reduktojnë urat disulfide shkakton zbërthimin e protomerëve dhe shkatërrimin e strukturës kuaternare të proteinës. Në tabelë 1 përmbledh të dhënat mbi lidhjet që stabilizojnë nivele të ndryshme të organizimit të molekulës së proteinës [shfaq] .
| Tabela 1. Karakteristikat e lidhjeve të përfshira në organizimin strukturor të proteinave | ||
| Niveli i organizimit | Llojet e lidhjeve (nga forca) | Lloji i komunikimit |
| Primar (zinxhir polipeptid linear) | Kovalente (e fortë) | Peptid - ndërmjet grupeve α-amino dhe α-karboksil të aminoacideve |
| Sekondare (a-spiralja, β-strukturat) | I dobët | Hidrogjeni - midis grupeve peptidike (çdo i pari dhe i katërti) i një zinxhiri polipeptid ose midis grupeve peptide të zinxhirëve polipeptidikë ngjitur |
| Kovalente (e fortë) | Disulfide - sythe disulfide brenda një rajoni linear të një zinxhiri polipeptid | |
| Terciar (globular, fibrilar) | Kovalente (e fortë) | Disulfid, izopeptid, ester - midis radikaleve anësore të aminoacideve të pjesëve të ndryshme të zinxhirit polipeptid |
| I dobët | Hidrogjeni - midis radikaleve anësore të aminoacideve të pjesëve të ndryshme të zinxhirit polipeptid Jonik (kripë) - midis grupeve të ngarkuara në mënyrë të kundërt të radikaleve anësore të aminoacideve të zinxhirit polipeptid Van der Waals - midis radikaleve anësore jopolare të aminoacideve të zinxhirit polipeptid |
|
| Kuaternar (globular, fibrilar) | I dobët | Jonik - midis grupeve të ngarkuara në mënyrë të kundërt të radikaleve anësore të aminoacideve të secilës prej nënnjësive Hidrogjeni - midis radikaleve anësore të mbetjeve të aminoacideve të vendosura në sipërfaqen e zonave kontaktuese të nënnjësive |
| Kovalente (e fortë) | Disulfidi - midis mbetjeve të cisteinës të secilës prej sipërfaqeve kontaktuese të nënnjësive të ndryshme | |
Veçoritë e organizimit strukturor të disa proteinave fibrilare
Organizimi strukturor i proteinave fibrilare ka një sërë veçorish në krahasim me proteinat globulare. Këto karakteristika mund të shihen në shembullin e keratinës, fibroinës dhe kolagjenit. Keratinat ekzistojnë në konformacione α- dhe β. α-keratinat dhe fibroina kanë një strukturë dytësore të palosur me shtresa, megjithatë, në keratin zinxhirët janë paralelë, dhe në fibroinë ato janë antiparalele (shih Fig. 3); Përveç kësaj, keratina përmban lidhje disulfide ndërzinxhirë, ndërsa fibroina nuk i ka ato. Thyerja e lidhjeve disulfide çon në ndarjen e zinxhirëve polipeptidikë në keratina. Përkundrazi, formimi i numrit maksimal të lidhjeve disulfide në keratina nëpërmjet ekspozimit ndaj agjentëve oksidues krijon një strukturë të fortë hapësinore. Në përgjithësi, në proteinat fibrilare, ndryshe nga ato globulare, ndonjëherë është e vështirë të bëhet dallimi rigoroz midis niveleve të ndryshme të organizimit. Nëse pranojmë (si për një proteinë globulare) që struktura terciare duhet të formohet duke vendosur një zinxhir polipeptid në hapësirë, dhe struktura kuaternare nga disa zinxhirë, atëherë në proteinat fibrilare disa zinxhirë polipeptidikë janë përfshirë tashmë në formimin e strukturës dytësore. . Një shembull tipik i një proteine fibrilare është kolagjeni, i cili është një nga proteinat më të bollshme në trupin e njeriut (rreth 1/3 e masës së të gjitha proteinave). Gjendet në inde që kanë forcë të lartë dhe shtrirje të ulët (kockat, tendinat, lëkurën, dhëmbët, etj.). Në kolagjen, një e treta e mbetjeve të aminoacideve janë glicina, dhe rreth një e katërta ose pak më shumë janë prolina ose hidroksiprolina.

Zinxhiri i izoluar polipeptid i kolagjenit (struktura primare) duket si një vijë e thyer. Ai përmban rreth 1000 aminoacide dhe ka një peshë molekulare prej rreth 10 5 (Fig. 5, a, b). Zinxhiri polipeptid është ndërtuar nga një treshe përsëritëse e aminoacideve (treshe) të përbërjes së mëposhtme: gly-A-B, ku A dhe B janë çdo aminoacid tjetër përveç glicinës (më shpesh prolina dhe hidroksiprolina). Zinxhirët polipeptidë të kolagjenit (ose zinxhirët α) gjatë formimit të strukturave dytësore dhe terciare (Fig. 5, c dhe d) nuk mund të prodhojnë α-spira tipike me simetri spirale. Prolina, hidroksiprolina dhe glicina (aminoacide antihelike) ndërhyjnë në këtë. Prandaj, tre zinxhirë α formojnë, si të thuash, spirale të përdredhura, si tre fije që mbështillen rreth një cilindri. Tre zinxhirë α spirale formojnë një strukturë kolagjeni të përsëritur të quajtur tropokolagjen (Fig. 5d). Tropokolagjeni në organizimin e tij është struktura terciare e kolagjenit. Unazat e sheshta të prolinës dhe hidroksiprolinës, të alternuara rregullisht përgjatë zinxhirit, i japin asaj ngurtësi, siç bëjnë lidhjet ndërzinxhirore midis zinxhirëve α të tropokolagjenit (kjo është arsyeja pse kolagjeni është rezistent ndaj shtrirjes). Tropokolagjeni është në thelb një nënnjësi e fibrileve të kolagjenit. Vendosja e nënnjësive të tropokolagjenit në strukturën kuaternare të kolagjenit ndodh në mënyrë hap pas hapi (Fig. 5e).
Stabilizimi i strukturave të kolagjenit ndodh për shkak të lidhjeve të ndërlidhura të hidrogjenit, jonike dhe van der Waals dhe një numri të vogël lidhjesh kovalente.
Zinxhirët α të kolagjenit kanë struktura të ndryshme kimike. Ekzistojnë lloje të ndryshme të zinxhirëve α 1 (I, II, III, IV) dhe zinxhirët α 2. Në varësi të cilat zinxhirë α 1 dhe α 2 përfshihen në formimin e spirales me tre fije të tropokolagjenit, dallohen katër lloje të kolagjenit:
- lloji i parë - dy α 1 (I) dhe një zinxhir α 2;
- lloji i dytë - tre zinxhirë α 1 (II);
- lloji i tretë - tre zinxhirë α 1 (III);
- lloji i katërt - tre zinxhirë α 1 (IV).
Kolagjeni më i zakonshëm është i llojit të parë: gjendet në indet e eshtrave, lëkurën, tendinat; kolagjeni i tipit 2 gjendet në indet e kërcit, etj. Një lloj indi mund të përmbajë lloje të ndryshme kolagjeni.
Grumbullimi i renditur i strukturave të kolagjenit, ngurtësia dhe inertiteti i tyre sigurojnë forcën e lartë të fibrave të kolagjenit. Proteinat e kolagjenit përmbajnë gjithashtu përbërës karbohidrate, pra janë komplekse protein-karbohidrate.
Kolagjeni është një proteinë jashtëqelizore që formohet nga qelizat e indit lidhës që gjenden në të gjitha organet. Prandaj, me dëmtimin e kolagjenit (ose ndërprerjen e formimit të tij), ndodhin shkelje të shumta të funksioneve mbështetëse të indit lidhës të organeve.
| Faqe 3 | faqet totale: 7 |
MODULI 1 STRUKTURA, VETITË DHE FUNKSIONET E PROTEINAVE
MODULI 1 STRUKTURA, VETITË DHE FUNKSIONET E PROTEINAVE
Struktura e modulit | Temat |
Njësia modulare 1 | 1.1. Organizimi strukturor i proteinave. Fazat e formimit të konformimit të proteinave vendase 1.2. Bazat e funksionimit të proteinave. Barnat si ligandë që ndikojnë në funksionin e proteinave 1.3. Denatyrimi i proteinave dhe mundësia e ripërtëritjes spontane të tyre |
Njësia modulare 2 | 1.4. Karakteristikat e strukturës dhe funksionimit të proteinave oligomerike duke përdorur shembullin e hemoglobinës 1.5. Ruajtja e konformacionit të proteinave vendase në kushte qelizore 1.6. Shumëllojshmëri proteinash. Familjet e proteinave duke përdorur shembullin e imunoglobulinave 1.7. Vetitë fiziko-kimike të proteinave dhe metodat e ndarjes së tyre |
Njësia modulare 1 ORGANIZIMI STRUKTUROR I PROTEINAVE MONOMERIKE DHE BAZAT E FUNKSIONIMIT TË TYRE
Objektivat e mësimit Të jetë i aftë:
1. Përdorni njohuri për veçoritë strukturore të proteinave dhe varësinë e funksioneve të proteinave nga struktura e tyre për të kuptuar mekanizmat e zhvillimit të proteinopative trashëgimore dhe të fituara.
2. Shpjegoni mekanizmat e veprimit terapeutik të disa barnave si ligandë që ndërveprojnë me proteinat dhe ndryshojnë aktivitetin e tyre.
3. Përdorni njohuri për strukturën dhe qëndrueshmërinë konformuese të proteinave për të kuptuar paqëndrueshmërinë e tyre strukturore dhe funksionale dhe tendencën për denatyrim në kushte të ndryshimit.
4. Shpjegoni përdorimin e agjentëve denatyrues si mjete për sterilizimin e materialeve dhe instrumenteve mjekësore, si dhe antiseptikëve.
Dije:
1. Nivelet e organizimit strukturor të proteinave.
2. Rëndësia e strukturës parësore të proteinave, e cila përcakton diversitetin strukturor dhe funksional të tyre.
3. Mekanizmi i formimit të qendrës aktive në proteina dhe ndërveprimi i saj specifik me ligand, i cili qëndron në themel të funksionimit të proteinave.
4. Shembuj të ndikimit të ligandëve ekzogjenë (barna, toksina, helme) në konformimin dhe aktivitetin funksional të proteinave.
5. Shkaqet dhe pasojat e denatyrimit të proteinave, faktorët që shkaktojnë denatyrimin.
6. Shembuj të përdorimit të faktorëve denatyrues në mjekësi si antiseptikë dhe mjete për sterilizimin e instrumenteve mjekësore.
TEMA 1.1. ORGANIZIMI STRUKTUROR I PROTEINAVE. FAZAT E FORMIMIT TË VENDES
KONFORMIMI I PROTEINËS
Proteinat janë molekula polimer, monomerët e të cilave janë vetëm 20 α-aminoacide. Seti dhe rendi i kombinimit të aminoacideve në një proteinë përcaktohet nga struktura e gjeneve në ADN-në e individëve. Çdo proteinë, në përputhje me strukturën e saj specifike, kryen funksionin e saj. Grupi i proteinave të një organizmi të caktuar përcakton karakteristikat e tij fenotipike, si dhe praninë e sëmundjeve trashëgimore ose një predispozicion për zhvillimin e tyre.
1. Aminoacidet që përbëjnë proteinat. Lidhja peptide. Proteinat janë polimere të ndërtuara nga monomere - 20 α-aminoacide, formula e përgjithshme e të cilave është
Aminoacidet ndryshojnë në strukturën, madhësinë dhe vetitë fiziko-kimike të radikalëve të lidhur me atomin α-karbon. Grupet funksionale të aminoacideve përcaktojnë karakteristikat e vetive të α-aminoacideve të ndryshme. Radikalet që gjenden në α-aminoacidet mund të ndahen në disa grupe:
Proline, Ndryshe nga 19 monomerët e tjerë të proteinave, ai nuk është një aminoacid, por një radikal në prolinë është i lidhur me atomin α-karbonit dhe me grupin imino;
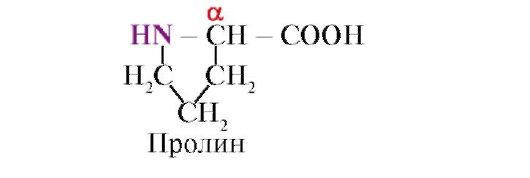 Aminoacidet ndryshojnë në tretshmërinë në ujë. Kjo është për shkak të aftësisë së radikalëve për të bashkëvepruar me ujin (hidrat).
Aminoacidet ndryshojnë në tretshmërinë në ujë. Kjo është për shkak të aftësisë së radikalëve për të bashkëvepruar me ujin (hidrat).
TE hidrofile përfshijnë radikale që përmbajnë grupe funksionale anionike, kationike dhe polare të pangarkuara.
TE hidrofobe përfshijnë radikale që përmbajnë grupe metil, zinxhirë alifatikë ose unaza.
2. Lidhjet peptide lidhin aminoacide për të formuar peptide. Gjatë sintezës së peptideve, grupi α-karboksil i një aminoacidi ndërvepron me grupin α-amino të një aminoacidi tjetër për të formuar lidhja peptide:
Proteinat janë polipeptide, d.m.th. polimere lineare të α-aminoacideve të lidhura nga një lidhje peptide (Fig. 1.1.)
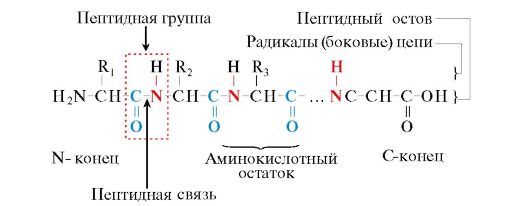 Oriz. 1.1. Termat e përdorur për të përshkruar strukturën e peptideve
Oriz. 1.1. Termat e përdorur për të përshkruar strukturën e peptideve
Monomerët e aminoacideve që përbëjnë polipeptidet quhen mbetjet e aminoacideve. Një zinxhir grupesh të përsëritura - NH-CH-CO- forma shtylla kurrizore peptide. Një mbetje e aminoacideve që ka një grup α-amino të lirë quhet N-terminal, dhe ai që ka një grup të lirë α-karboksil quhet C-terminal. Peptidet shkruhen dhe lexohen nga fundi N në fundin C.
Lidhja peptide e formuar nga grupi imino e prolinës ndryshon nga lidhjet e tjera peptide: atomit të azotit të grupit peptid i mungon hidrogjeni,
në vend të kësaj, ekziston një lidhje me një radikal, si rezultat i së cilës njëra anë e unazës përfshihet në shtyllën kurrizore peptide:
 Peptidet ndryshojnë në përbërjen e aminoacideve, numrin e aminoacideve dhe renditjen e lidhjes së aminoacideve, për shembull, Ser-Ala-Glu-Gis dhe His-Glu-Ala-Ser janë dy peptide të ndryshme.
Peptidet ndryshojnë në përbërjen e aminoacideve, numrin e aminoacideve dhe renditjen e lidhjes së aminoacideve, për shembull, Ser-Ala-Glu-Gis dhe His-Glu-Ala-Ser janë dy peptide të ndryshme.
Lidhjet peptide janë shumë të forta dhe hidroliza e tyre kimike jo enzimatike kërkon kushte të vështira: proteina që analizohet hidrolizohet në acid klorhidrik të përqendruar në një temperaturë prej rreth 110° për 24 orë. Në një qelizë të gjallë, lidhjet peptide mund të thyhen enzimat proteolitike, thirrur proteazat ose hidrolaza peptide.
3. Struktura primare e proteinave. Mbetjet e aminoacideve në zinxhirët peptidikë të proteinave të ndryshme nuk alternohen rastësisht, por janë të renditura në një rend të caktuar. Sekuenca lineare ose rendi i alternimit i mbetjeve të aminoacideve në një zinxhir polipeptid quhet struktura primare e proteinës.
Struktura primare e secilës proteinë individuale është e koduar në molekulën e ADN-së (në një rajon të quajtur gjen) dhe realizohet gjatë transkriptimit (kopjimi i informacionit në mARN) dhe përkthimi (sinteza e strukturës primare të proteinës). Rrjedhimisht, struktura primare e proteinave të një personi individual është informacioni i transmetuar trashëgimisht nga prindërit te fëmijët, i cili përcakton veçoritë strukturore të proteinave të një organizmi të caktuar, nga të cilat varet funksioni i proteinave ekzistuese (Fig. 1.2.).
 Oriz. 1.2. Marrëdhënia midis gjenotipit dhe konformacionit të proteinave të sintetizuara në trupin e individit
Oriz. 1.2. Marrëdhënia midis gjenotipit dhe konformacionit të proteinave të sintetizuara në trupin e individit
Secila prej rreth 100,000 proteinave individuale në trupin e njeriut ka unike struktura primare. Molekulat e të njëjtit lloj proteine (për shembull, albumina) kanë të njëjtin alternim të mbetjeve të aminoacideve, gjë që e dallon albuminën nga çdo proteinë tjetër individuale.
Sekuenca e mbetjeve të aminoacideve në një zinxhir peptid mund të konsiderohet një formë e regjistrimit të informacionit. Ky informacion përcakton rregullimin hapësinor të zinxhirit linear peptid në një strukturë më kompakte tre-dimensionale të quajtur konformacion ketri. Procesi i formimit të një konformacioni proteinik funksionalisht aktiv quhet palosshme
4. Konformimi i proteinave. Rrotullimi i lirë në shtyllën kurrizore peptide është i mundur midis atomit të azotit të grupit peptid dhe atomit fqinj α-karbonit, si dhe midis atomit të α-karbonit dhe karbonit të grupit karbonil. Për shkak të ndërveprimit të grupeve funksionale të mbetjeve të aminoacideve, struktura primare e proteinave mund të fitojë struktura hapësinore më komplekse. Në proteinat globulare, ekzistojnë dy nivele kryesore të palosjes së konformacionit të zinxhirëve peptidikë: dytësore Dhe strukturë terciare.
Struktura dytësore e proteinaveështë një strukturë hapësinore e formuar si rezultat i formimit të lidhjeve hidrogjenore midis grupeve funksionale -C=O dhe -NH- të shtyllës kurrizore peptide. Në këtë rast, zinxhiri peptid mund të fitojë struktura të rregullta të dy llojeve: α-helika Dhe β-strukturat.
NË α-helika lidhjet hidrogjenore formohen midis atomit të oksigjenit të grupit karbonil dhe hidrogjenit të azotit amid të aminoacidit të 4-të; zinxhirët anësor të mbetjeve të aminoacideve
ndodhen përgjatë periferisë së spirales, pa marrë pjesë në formimin e strukturës dytësore (Fig. 1.3.).
Radikalët e shumtë, ose radikalët që mbajnë ngarkesa të barabarta, parandalojnë formimin e një spirale α. Mbetja e prolinës, e cila ka një strukturë unazore, ndërpret heliksin α, pasi për shkak të mungesës së hidrogjenit në atomin e azotit në zinxhirin peptid është e pamundur të formohet një lidhje hidrogjeni. Lidhja midis azotit dhe atomit të karbonit α është pjesë e unazës së prolinës, kështu që shtylla kurrizore peptide bëhet e përkulur në këtë pikë.
β-Strukturaështë formuar midis rajoneve lineare të shtyllës kurrizore peptide të një zinxhiri polipeptid, duke formuar kështu struktura të palosura. Mund të formohen zinxhirë polipeptidikë ose pjesë të tyre paralele ose β-struktura antiparalele. Në rastin e parë, skajet N dhe C të vargjeve peptide ndërvepruese përputhen dhe në të dytën kanë drejtim të kundërt (Fig. 1.4).
Oriz. 1.3. Struktura dytësore e proteinave - α-helix
 Oriz. 1.4. Strukturat e fletës β paralele dhe antiparalele
Oriz. 1.4. Strukturat e fletës β paralele dhe antiparalele
Strukturat β tregohen me shigjeta të gjera: A - Struktura β antiparalele. B - Strukturat paralele të fletës β
Në disa proteina, strukturat β mund të formohen për shkak të formimit të lidhjeve hidrogjenore midis atomeve të shtyllës kurrizore peptide të zinxhirëve të ndryshëm polipeptidikë.
Gjendet edhe në proteina zona me sekondare të parregullta strukturë, e cila përfshin kthesat, sythe dhe kthesat e shtyllës kurrizore të polipeptidit. Ato shpesh ndodhen në vende ku drejtimi i zinxhirit peptid ndryshon, për shembull, kur formohet një strukturë paralele e fletës β.
Bazuar në praninë e α-helikave dhe strukturave β, proteinat globulare mund të ndahen në katër kategori.
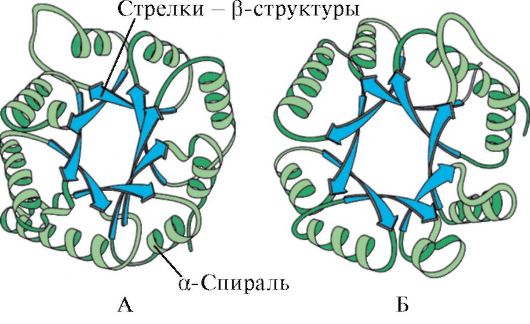
Oriz. 1.5. Struktura dytësore e mioglobinës (A) dhe zinxhirit β të hemoglobinës (B), që përmban tetë α-helika

 Oriz. 1.6. Struktura dytësore e domenit të izomerazës triosefosfat dhe piruvat kinazës
Oriz. 1.6. Struktura dytësore e domenit të izomerazës triosefosfat dhe piruvat kinazës
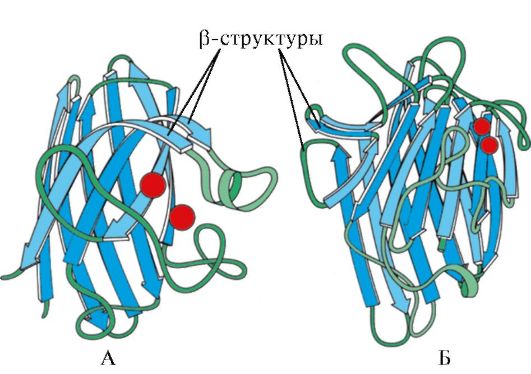 Oriz. 1.7. Struktura dytësore e domenit konstant të imunoglobulinës (A) dhe enzimës superoksid dismutazë (B)
Oriz. 1.7. Struktura dytësore e domenit konstant të imunoglobulinës (A) dhe enzimës superoksid dismutazë (B)
NË kategoria e katërt përfshinte proteina që përmbajnë një sasi të vogël të strukturave të rregullta dytësore. Këto proteina përfshijnë proteina të vogla të pasura me cisteinë ose metaloproteina.
Struktura terciare e proteinave- një lloj konformacioni i formuar për shkak të ndërveprimeve midis radikaleve të aminoacideve, të cilat mund të vendosen në një distancë të konsiderueshme nga njëri-tjetri në zinxhirin peptid. Shumica e proteinave formojnë një strukturë hapësinore që i ngjan një rruzulli (proteina globulare).
Meqenëse radikalet hidrofobike të aminoacideve priren të kombinohen përmes të ashtuquajturave ndërveprimet hidrofobike dhe forcat ndërmolekulare të van der Waals-it, një bërthamë e dendur hidrofobike formohet brenda globulës së proteinave. Radikalet hidrofile të jonizuara dhe jojonizuese janë kryesisht të vendosura në sipërfaqen e proteinës dhe përcaktojnë tretshmërinë e saj në ujë.
 Oriz. 1.8. Llojet e lidhjeve që lindin midis radikaleve të aminoacideve gjatë formimit të strukturës terciare të një proteine
Oriz. 1.8. Llojet e lidhjeve që lindin midis radikaleve të aminoacideve gjatë formimit të strukturës terciare të një proteine
1 - lidhje jonike- ndodh ndërmjet grupeve funksionale të ngarkuara pozitivisht dhe negativisht;
2 - lidhje hidrogjenore- ndodh ndërmjet një grupi hidrofil të pa ngarkuar dhe çdo grupi tjetër hidrofil;
3 - ndërveprimet hidrofobike- lindin midis radikaleve hidrofobe;
4 - lidhje disulfide- formuar për shkak të oksidimit të grupeve SH të mbetjeve të cisteinës dhe ndërveprimit të tyre me njëri-tjetrin
Mbetjet hidrofile të aminoacideve që gjenden brenda bërthamës hidrofobike mund të ndërveprojnë me njëra-tjetrën duke përdorur jonike Dhe lidhjet hidrogjenore(Fig. 1.8).
Lidhjet jonike dhe hidrogjenore, si dhe ndërveprimet hidrofobike, janë të dobëta: energjia e tyre nuk është shumë më e lartë se energjia e lëvizjes termike të molekulave në temperaturën e dhomës. Konformimi i proteinës ruhet nga formimi i shumë lidhjeve të tilla të dobëta. Meqenëse atomet që përbëjnë një proteinë janë në lëvizje të vazhdueshme, është e mundur të thyhen disa lidhje të dobëta dhe të formohen të tjera, gjë që çon në lëvizje të lehta të seksioneve individuale të zinxhirit polipeptid. Kjo veti e proteinave për të ndryshuar konformimin si rezultat i thyerjes së disa dhe formimit të lidhjeve të tjera të dobëta quhet qëndrueshmëri konformuese.
Trupi i njeriut ka sisteme që mbështesin homeostaza- qëndrueshmëria e mjedisit të brendshëm brenda kufijve të caktuar të pranueshëm për një trup të shëndetshëm. Në kushtet e homeostazës, ndryshimet e vogla në konformacion nuk e prishin strukturën dhe funksionin e përgjithshëm të proteinave. Konformacioni funksionalisht aktiv i një proteine quhet konformacion vendas. Ndryshimet në mjedisin e brendshëm (për shembull, përqendrimi i glukozës, joneve të Ca, protoneve, etj.) çojnë në ndryshime në konformimin dhe ndërprerje të funksioneve të proteinave.
Struktura terciare e disa proteinave është e stabilizuar lidhjet disulfide, e formuar për shkak të bashkëveprimit të -SH grupeve të dy mbetjeve
 Oriz. 1.9. Formimi i një lidhje disulfide në një molekulë proteine
Oriz. 1.9. Formimi i një lidhje disulfide në një molekulë proteine
cisteinë (Fig. 1.9). Shumica e proteinave ndërqelizore nuk kanë lidhje kovalente disulfide në strukturën e tyre terciare. Prania e tyre është karakteristikë e proteinave të sekretuara nga qeliza, gjë që siguron stabilitet më të madh të tyre në kushte jashtëqelizore. Kështu, lidhjet disulfide janë të pranishme në molekulat e insulinës dhe imunoglobulinave.
Insulinë- një hormon proteinik i sintetizuar në qelizat β të pankreasit dhe i sekretuar në gjak në përgjigje të rritjes së përqendrimit të glukozës në gjak.
 Në strukturën e insulinës, ekzistojnë dy lidhje disulfide që lidhin zinxhirët polipeptid A dhe B, dhe një lidhje disulfide brenda zinxhirit A (Fig. 1.10).
Në strukturën e insulinës, ekzistojnë dy lidhje disulfide që lidhin zinxhirët polipeptid A dhe B, dhe një lidhje disulfide brenda zinxhirit A (Fig. 1.10).
Oriz. 1.10. Lidhjet disulfide në strukturën e insulinës 5. Struktura supersekondare e proteinave. Në proteinat me strukturë dhe funksione të ndryshme parësore, ato ndonjëherë zbulohen kombinime të ngjashme dhe pozicione relative të strukturave dytësore,
të cilat quhen strukturë supersekondare. Ai zë një pozicion të ndërmjetëm midis strukturave dytësore dhe terciare, pasi është një kombinim specifik i elementeve të strukturës sekondare në formimin e strukturës terciare të proteinës. Strukturat supersekondare kanë emra specifikë, të tillë si "a-helix-turn-a-helix", "zinxher leucine", "zink gishtat", etj. Struktura të tilla supersekondare janë karakteristike për proteinat që lidhen me ADN-në."Zinxhir leucine".
 Ky lloj strukture supersekondare përdoret për të bashkuar dy proteina së bashku. Në sipërfaqen e proteinave ndërvepruese ka zona α-spiralike që përmbajnë të paktën katër mbetje leucine. Mbetjet e leucinës në α-helix janë të vendosura gjashtë aminoacide larg njëri-tjetrit. Meqenëse çdo kthesë e heliksit α përmban 3.6 mbetje aminoacide, radikalet e leucinës ndodhen në sipërfaqen e çdo kthese të dytë. Mbetjet e leucinës së heliksit α të një proteine mund të ndërveprojnë me mbetjet e leucinës të një proteine tjetër (ndërveprime hidrofobike), duke i lidhur ato së bashku (Fig. 1.11.). Shumë proteina lidhëse të ADN-së funksionojnë në komplekset oligomerike ku nën-njësitë individuale janë të lidhura me njëra-tjetrën me "zinxkë leucine".
Ky lloj strukture supersekondare përdoret për të bashkuar dy proteina së bashku. Në sipërfaqen e proteinave ndërvepruese ka zona α-spiralike që përmbajnë të paktën katër mbetje leucine. Mbetjet e leucinës në α-helix janë të vendosura gjashtë aminoacide larg njëri-tjetrit. Meqenëse çdo kthesë e heliksit α përmban 3.6 mbetje aminoacide, radikalet e leucinës ndodhen në sipërfaqen e çdo kthese të dytë. Mbetjet e leucinës së heliksit α të një proteine mund të ndërveprojnë me mbetjet e leucinës të një proteine tjetër (ndërveprime hidrofobike), duke i lidhur ato së bashku (Fig. 1.11.). Shumë proteina lidhëse të ADN-së funksionojnë në komplekset oligomerike ku nën-njësitë individuale janë të lidhura me njëra-tjetrën me "zinxkë leucine".
Oriz. 1.11. "Zinxhir leucine" midis zonave α-spiral të dy proteinave Një shembull i proteinave të tilla janë histonet.- proteinat bërthamore, të cilat përmbajnë një numër të madh të aminoacideve të ngarkuar pozitivisht - argininë dhe lizinë (deri në 80%). Molekulat e histonit kombinohen në komplekse oligomerike që përmbajnë tetë monomere duke përdorur "zippers leucine", pavarësisht ngarkesës domethënëse homonime të këtyre molekulave.
"Gishti i zinkut"- një variant i strukturës supersekondare, karakteristik për proteinat që lidhin ADN-në, ka formën e një fragmenti të zgjatur në sipërfaqen e proteinës dhe përmban rreth 20 mbetje aminoacide (Fig. 1.12). Forma e "gishtit të zgjatur" mbështetet nga një atom zinku i lidhur me katër radikale aminoacide - dy mbetje cisteine dhe dy mbetje histidine. Në disa raste, në vend të mbetjeve të histidinës, ka mbetje të cisteinës. Dy mbetje cisteine të vendosura afër ndahen nga dy mbetjet e tjera Gisili nga një sekuencë Cys e përbërë nga afërsisht 12 mbetje aminoacide. Ky rajon i proteinës formon një α-helix, radikalët e së cilës mund të lidhen në mënyrë specifike me rajonet rregulluese të brazdës kryesore të ADN-së. Specifikimi i lidhjes individuale
 Oriz. 1.12. Struktura primare e rajonit të proteinave që lidhin ADN-në që formojnë strukturën e "gishtit të zinkut" (shkronjat tregojnë aminoacidet që përbëjnë këtë strukturë)
Oriz. 1.12. Struktura primare e rajonit të proteinave që lidhin ADN-në që formojnë strukturën e "gishtit të zinkut" (shkronjat tregojnë aminoacidet që përbëjnë këtë strukturë)
Proteina rregullatore e lidhjes së ADN-së varet nga sekuenca e mbetjeve të aminoacideve të vendosura në rajonin e gishtit të zinkut. Struktura të tilla përmbajnë, në veçanti, receptorë për hormonet steroide të përfshirë në rregullimin e transkriptimit (leximi i informacionit nga ADN në ARN).
TEMA 1.2. BAZAT E FUNKSIONIMIT TË PROTEINAVE. BARNAT SI LIGANDË QË NDIKOJNË NË FUNKSIONIN E PROTEINËS
1. Qendra aktive e proteinës dhe ndërveprimi i saj me ligand. Gjatë formimit të strukturës terciare, një rajon formohet në sipërfaqen e një proteine funksionalisht aktive, zakonisht në një gropë, të formuar nga radikale aminoacide që janë larg njëri-tjetrit në strukturën parësore. Ky rajon, i cili ka një strukturë unike për një proteinë të caktuar dhe është i aftë të ndërveprojë në mënyrë specifike me një molekulë të veçantë ose grup molekulash të ngjashme, quhet vendi i lidhjes protein-ligand ose zona aktive. Ligandët janë molekula që ndërveprojnë me proteinat.
Specifikimi i lartë Ndërveprimi i proteinës me ligand sigurohet nga komplementariteti i strukturës së qendrës aktive me strukturën e ligandit.
Komplementariteti- kjo është korrespondenca hapësinore dhe kimike e sipërfaqeve ndërvepruese. Qendra aktive jo vetëm që duhet të korrespondojë hapësinore me ligandin e përfshirë në të, por edhe lidhjet (ndërveprimet jonike, hidrogjeni dhe hidrofobike) duhet të formohen midis grupeve funksionale të radikalëve të përfshirë në qendrën aktive dhe ligandit që mbajnë ligand. në qendrën aktive (Fig. 1.13 ).
 Oriz. 1.13. Ndërveprimi plotësues i proteinës me ligand
Oriz. 1.13. Ndërveprimi plotësues i proteinës me ligand
Disa ligandë, kur lidhen me qendrën aktive të një proteine, luajnë një rol ndihmës në funksionimin e proteinave. Ligandë të tillë quhen kofaktorë, dhe proteinat që përmbajnë një pjesë jo proteinike quhen proteinat komplekse(në krahasim me proteinat e thjeshta, që përbëhen vetëm nga pjesa proteinike). Pjesa jo proteinike, e lidhur fort me proteinën, quhet grup protetik. Për shembull, mioglobina, hemoglobina dhe citokromet përmbajnë një grup protetik, hemin, që përmban një jon hekuri, të lidhur fort me qendrën aktive. Proteinat komplekse që përmbajnë hem quhen hemoproteina.
Kur ligandët specifikë janë të lidhur me proteinat, funksioni i këtyre proteinave manifestohet. Kështu, albumina, proteina më e rëndësishme në plazmën e gjakut, shfaq funksionin e saj transportues duke bashkangjitur ligandët hidrofobikë, si acidet yndyrore, bilirubina, disa ilaçe, etj. në qendrën aktive (Fig. 1.14).
Ligandët që ndërveprojnë me strukturën tredimensionale të zinxhirit peptid mund të jenë jo vetëm molekula organike dhe inorganike me molekula të ulëta, por edhe makromolekula:
ADN (shembuj me proteinat që lidhin ADN-në të diskutuar më sipër);
Polisakaridet;
 Oriz. 1.14. Marrëdhënia midis gjenotipit dhe fenotipit
Oriz. 1.14. Marrëdhënia midis gjenotipit dhe fenotipit
Struktura unike parësore e proteinave njerëzore, e koduar në molekulën e ADN-së, realizohet në qeliza në formën e një konformimi unik, strukturë qendrore aktive dhe funksione proteinike.
Në këto raste, proteina njeh një rajon specifik të ligandit që është proporcional dhe plotësues me vendin e lidhjes. Kështu, në sipërfaqen e hepatociteve ka proteina receptore për hormonin insulinë, i cili gjithashtu ka një strukturë proteinike. Ndërveprimi i insulinës me receptorin shkakton një ndryshim në konformimin e tij dhe aktivizimin e sistemeve të sinjalizimit, duke çuar në ruajtjen e lëndëve ushqyese në hepatocitet pas ngrënies.
Kështu, Funksionimi i proteinave bazohet në ndërveprimin specifik të qendrës aktive të proteinës me ligand.
2. Struktura e domenit dhe roli i tij në funksionimin e proteinave. Zinxhirët e gjatë polipeptidikë të proteinave globulare shpesh palosen në disa rajone kompakte, relativisht të pavarura. Ato kanë një strukturë terciare të pavarur, që të kujton atë të proteinave globulare dhe quhen domenet. Për shkak të strukturës së domenit të proteinave, struktura e tyre terciare është më e lehtë për t'u formuar.
Në proteinat e domenit, vendet e lidhjes së ligandit shpesh ndodhen midis domeneve. Kështu, tripsina është një enzimë proteolitike që prodhohet nga pjesa ekzokrine e pankreasit dhe është e nevojshme për tretjen e proteinave ushqimore. Ajo ka një strukturë me dy domene, dhe qendra e lidhjes së tripsinës me ligandin e saj - proteinën ushqimore - ndodhet në brazdë midis dy domeneve. Në qendrën aktive krijohen kushtet e nevojshme për lidhjen efektive të një vendi specifik të proteinës ushqimore dhe hidrolizën e lidhjeve të saj peptide.
Domenet e ndryshme në një proteinë mund të lëvizin në lidhje me njëra-tjetrën kur qendra aktive ndërvepron me ligand (Fig. 1.15).
Heksokinaza- një enzimë që katalizon fosforilimin e glukozës duke përdorur ATP. Vendi aktiv i enzimës ndodhet në çarjen midis dy domeneve. Kur heksokinaza lidhet me glukozën, domenet që e rrethojnë mbyllen dhe substrati bllokohet, ku ndodh fosforilimi (shih Fig. 1.15).
 Oriz. 1.15. Lidhja e domeneve të heksokinazës me glukozën
Oriz. 1.15. Lidhja e domeneve të heksokinazës me glukozën
Në disa proteina, domenet kryejnë funksione të pavarura duke u lidhur me ligandë të ndryshëm. Proteinat e tilla quhen multifunksionale.
3. Barnat janë ligandë që ndikojnë në funksionin e proteinave. Ndërveprimi i proteinave me ligandët është specifik. Megjithatë, për shkak të qëndrueshmërisë konformuese të proteinës dhe qendrës së saj aktive, është e mundur të zgjidhet një substancë tjetër që gjithashtu mund të ndërveprojë me proteinën në qendrën aktive ose një pjesë tjetër të molekulës.
Një substancë e ngjashme në strukturë me një ligand natyror quhet analog strukturor i ligandit ose një ligand jo-natyror. Ai gjithashtu ndërvepron me proteinën në vendin aktiv. Një analog strukturor i një ligandi mund të përmirësojë funksionin e proteinave (agonist), dhe zvogëloni atë (antagonist). Ligandi dhe analogët e tij strukturorë konkurrojnë me njëri-tjetrin për t'u lidhur me proteinën në të njëjtin vend. Substancat e tilla quhen modulatorët konkurrues(rregullatorë) të funksioneve të proteinave. Shumë ilaçe veprojnë si frenues të proteinave. Disa prej tyre përftohen nga modifikimi kimik i ligandëve natyrorë. Frenuesit e funksioneve të proteinave mund të jenë ilaçet dhe helmet.
Atropina është një frenues konkurrues i receptorëve M-kolinergjikë. Acetilkolina është një neurotransmetues për transmetimin e impulseve nervore përmes sinapseve kolinergjike. Për të kryer ngacmimin, acetilkolina e lëshuar në çarjen sinaptike duhet të ndërveprojë me proteinën e receptorit të membranës postsinaptike. Dy lloje të gjetura receptorët kolinergjikë:
M receptor përveç acetilkolinës, ndërvepron në mënyrë selektive me muskarinën (toksina agarike e mizës). M - receptorët kolinergjikë janë të pranishëm në muskujt e lëmuar dhe, kur ndërveprojnë me acetilkolinën, shkaktojnë tkurrjen e tyre;
receptori H në mënyrë specifike lidhet me nikotinën. Receptorët N-kolinergjikë gjenden në sinapset e muskujve skeletorë të strijuar.
Frenues specifik Receptorët M-kolinergjikëështë atropinë. Gjendet në bimët e belladonës dhe pulës.
 Atropina ka grupe funksionale të ngjashme në strukturë me acetilkolinën dhe rregullimin e tyre hapësinor, prandaj është një frenues konkurrues i receptorëve M-kolinergjikë. Duke marrë parasysh që lidhja e acetilkolinës me receptorët M-kolinergjikë shkakton tkurrje të muskujve të lëmuar, atropina përdoret si ilaç që lehtëson spazmën e tyre. (antispazmatike). Kështu, dihet përdorimi i atropinës për të relaksuar muskujt e syrit gjatë shikimit të fundusit, si dhe për të lehtësuar spazmat gjatë dhimbjeve të barkut gastrointestinal. Receptorët M-kolinergjikë janë të pranishëm edhe në sistemin nervor qendror (SNQ), kështu që doza të mëdha të atropinës mund të shkaktojnë një reagim të padëshiruar nga sistemi nervor qendror: agjitacion motorik dhe mendor, halucinacione, konvulsione.
Atropina ka grupe funksionale të ngjashme në strukturë me acetilkolinën dhe rregullimin e tyre hapësinor, prandaj është një frenues konkurrues i receptorëve M-kolinergjikë. Duke marrë parasysh që lidhja e acetilkolinës me receptorët M-kolinergjikë shkakton tkurrje të muskujve të lëmuar, atropina përdoret si ilaç që lehtëson spazmën e tyre. (antispazmatike). Kështu, dihet përdorimi i atropinës për të relaksuar muskujt e syrit gjatë shikimit të fundusit, si dhe për të lehtësuar spazmat gjatë dhimbjeve të barkut gastrointestinal. Receptorët M-kolinergjikë janë të pranishëm edhe në sistemin nervor qendror (SNQ), kështu që doza të mëdha të atropinës mund të shkaktojnë një reagim të padëshiruar nga sistemi nervor qendror: agjitacion motorik dhe mendor, halucinacione, konvulsione.
Ditilin është një agonist konkurrues i receptorëve H-kolinergjikë, duke frenuar funksionin e sinapsave neuromuskulare.
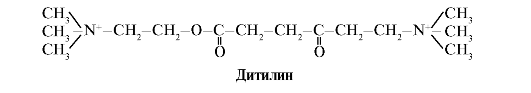 Sinapsat neuromuskulare të muskujve skeletorë përmbajnë receptorë H-kolinergjikë. Ndërveprimi i tyre me acetilkolinën çon në tkurrje të muskujve. Gjatë disa operacioneve kirurgjikale, si dhe në studimet endoskopike, përdoren barna që shkaktojnë relaksim të muskujve skeletorë. (relaksues të muskujve). Këto përfshijnë ditilinën, e cila është një analog strukturor i acetilkolinës. Ai lidhet me receptorët H-kolinergjikë, por ndryshe nga acetilkolina, ajo shkatërrohet shumë ngadalë nga enzima acetilkolinesterazë. Si rezultat i hapjes së zgjatur të kanaleve jonike dhe depolarizimit të vazhdueshëm të membranës, përçimi i impulseve nervore prishet dhe ndodh relaksimi i muskujve. Fillimisht, këto veti u zbuluan në helmin curare, për këtë arsye quhen ilaçe të tilla si kurare.
Sinapsat neuromuskulare të muskujve skeletorë përmbajnë receptorë H-kolinergjikë. Ndërveprimi i tyre me acetilkolinën çon në tkurrje të muskujve. Gjatë disa operacioneve kirurgjikale, si dhe në studimet endoskopike, përdoren barna që shkaktojnë relaksim të muskujve skeletorë. (relaksues të muskujve). Këto përfshijnë ditilinën, e cila është një analog strukturor i acetilkolinës. Ai lidhet me receptorët H-kolinergjikë, por ndryshe nga acetilkolina, ajo shkatërrohet shumë ngadalë nga enzima acetilkolinesterazë. Si rezultat i hapjes së zgjatur të kanaleve jonike dhe depolarizimit të vazhdueshëm të membranës, përçimi i impulseve nervore prishet dhe ndodh relaksimi i muskujve. Fillimisht, këto veti u zbuluan në helmin curare, për këtë arsye quhen ilaçe të tilla si kurare.
TEMA 1.3. DENATURIMI I PROTEINAVE DHE MUNDËSIA E RINATIVIMIT SPONTANE TË TYRE
1. Meqenëse konformacioni vendas i proteinave ruhet për shkak të ndërveprimeve të dobëta, ndryshimet në përbërjen dhe vetitë e mjedisit që rrethon proteinën, ekspozimi ndaj reagentëve kimikë dhe faktorëve fizikë shkaktojnë një ndryshim në konformimin e tyre (vetia e qëndrueshmërisë konformuese). Thyerja e një numri të madh lidhjesh çon në shkatërrimin e konformimit vendas dhe denatyrimin e proteinave.
Denatyrimi i proteinave- ky është shkatërrimi i konformacionit të tyre vendas nën ndikimin e agjentëve denatyrues, i shkaktuar nga këputja e lidhjeve të dobëta që stabilizojnë strukturën hapësinore të proteinës. Denatyrimi shoqërohet me shkatërrimin e strukturës unike tre-dimensionale dhe qendrës aktive të proteinës dhe humbjen e aktivitetit të saj biologjik (Fig. 1.16).
Të gjitha molekulat e denatyruara të një proteine fitojnë një konformacion të rastësishëm që ndryshon nga molekulat e tjera të së njëjtës proteinë. Radikalet e aminoacideve që formojnë qendrën aktive rezultojnë të jenë të largët hapësinor nga njëri-tjetri, d.m.th. vendi specifik i lidhjes së proteinës me ligand është shkatërruar. Gjatë denatyrimit, struktura primare e proteinave mbetet e pandryshuar.
Aplikimi i agjentëve denatyrues në kërkimin biologjik dhe mjekësi. Në studimet biokimike, përpara se të përcaktohen komponimet me peshë të ulët molekulare në materialin biologjik, proteinat zakonisht hiqen fillimisht nga tretësira. Acidi trikloroacetik (TCA) përdoret më shpesh për këtë qëllim. Pas shtimit të TCA në tretësirë, proteinat e denatyruara precipitojnë dhe hiqen lehtësisht me filtrim (Tabela 1.1.)
Në mjekësi, agjentët denatyrues shpesh përdoren për të sterilizuar instrumentet dhe materialet mjekësore në autoklava (agjenti denatyrues është temperatura e lartë) dhe si antiseptikë (alkool, fenol, kloraminë) për të trajtuar sipërfaqet e kontaminuara që përmbajnë mikroflora patogjene.
2. Riaktivizimi spontan i proteinave- vërtetimi i determinizmit të strukturës parësore, konformimit dhe funksionit të proteinave. Proteinat individuale janë produkte të një gjeni që kanë një sekuencë identike të aminoacideve dhe fitojnë të njëjtin konformacion në qelizë. Konkluzioni themelor se struktura primare e një proteine tashmë përmban informacion në lidhje me konformimin dhe funksionin e saj u bë në bazë të aftësisë së disa proteinave (në veçanti, ribonukleazës dhe mioglobinës) për t'u ripërtërirë spontanisht - për të rivendosur konformitetin e tyre amtare pas denatyrimit.
Formimi i strukturave të proteinave hapësinore kryhet me metodën e vetë-montimit - një proces spontan në të cilin një zinxhir polipeptid, i cili ka një strukturë primare unike, tenton të adoptojë një konformacion me energjinë më të ulët të lirë në tretësirë. Aftësia për të ripërtërirë proteinat që ruajnë strukturën e tyre primare pas denatyrimit u përshkrua në një eksperiment me enzimën ribonukleazë.
Ribonukleaza është një enzimë që prish lidhjet midis nukleotideve individuale në një molekulë ARN. Kjo proteinë globulare ka një zinxhir polipeptid, struktura terciare e të cilit stabilizohet nga shumë lidhje të dobëta dhe katër lidhje disulfide.
Trajtimi i ribonukleazës me ure, e cila thyen lidhjet hidrogjenore në molekulë, dhe një agjent reduktues, i cili thyen lidhjet disulfide, çon në denatyrim të enzimës dhe humbje të aktivitetit të saj.
Heqja e agjentëve denatyrues me dializë çon në rivendosjen e konformacionit dhe funksionit të proteinave, d.m.th. te rilindja. (Fig. 1.17).
 Oriz. 1.17. Denatyrimi dhe renativimi i ribonukleazës
Oriz. 1.17. Denatyrimi dhe renativimi i ribonukleazës
A - konformacion vendas i ribonukleazës, në strukturën terciare të së cilës ka katër lidhje disulfide; B - molekula e ribonukleazës së denatyruar;
B - molekula e ribonukleazës e riaktivizuar me strukturë dhe funksion të rivendosur
1. Plotësoni tabelën 1.2.
Tabela 1.2. Klasifikimi i aminoacideve sipas polaritetit të radikaleve
2. Shkruani formulën e tetrapeptidit:
Asp - Pro - Fen - Liz
a) nënvizoni grupet përsëritëse në peptid që formojnë shtyllën kurrizore peptide dhe grupet e ndryshueshme të përfaqësuara nga radikalet e aminoacideve;
b) emërtoni skajet N dhe C;
c) nxjerr në pah lidhjet peptide;
d) shkruani një peptid tjetër që përbëhet nga të njëjtat aminoacide;
e) numëroni numrin e varianteve të mundshme të një tetrapeptidi me përbërje të ngjashme aminoacide.
3. Shpjegoni rolin e strukturës primare të proteinave duke përdorur shembullin e një analize krahasuese të dy hormoneve peptide strukturore të ngjashme dhe evolucionarisht të afërta të neurohipofizës së gjitarëve - oksitocina dhe vazopresina (Tabela 1.3).
Tabela 1.3. Struktura dhe funksionet e oksitocinës dhe vazopresinës
 Për ta bërë këtë:
Për ta bërë këtë:
a) krahasoni përbërjen dhe sekuencën e aminoacideve të dy peptideve;
b) të gjejë ngjashmërinë e strukturës parësore të dy peptideve dhe ngjashmërinë e veprimit të tyre biologjik;
c) të gjejë dallimet në strukturën e dy peptideve dhe dallimet në funksionet e tyre;
d) nxirret një përfundim për ndikimin e strukturës parësore të peptideve në funksionet e tyre.
4. Përshkruani fazat kryesore të formimit të konformimit të proteinave globulare (strukturat dytësore, terciare, koncepti i strukturës supersekondare). Tregoni llojet e lidhjeve të përfshira në formimin e strukturave proteinike. Cilat radikale aminoacide mund të marrin pjesë në formimin e ndërveprimeve hidrofobike, lidhjeve jonike dhe hidrogjenore.
Jepni shembuj.
5. Përcaktoni konceptin e "qëndrueshmërisë konformuese të proteinave", tregoni arsyet e ekzistencës dhe rëndësisë së tij.
6. Zgjeroni kuptimin e frazës së mëposhtme: "Funksionimi i proteinave bazohet në ndërveprimin e tyre specifik me ligand", duke përdorur termat dhe duke shpjeguar kuptimin e tyre: konformimi i proteinave, qendra aktive, ligandi, komplementariteti, funksioni i proteinave.
7. Duke përdorur një shembull, shpjegoni se cilat janë domenet dhe cili është roli i tyre në funksionimin e proteinave.
DETYRAT E VETËKONTROLLIT
1. Ndeshje.
Grupi funksional në radikal aminoacid:
A. Grupi karboksil B. Grupi hidroksil C Grupi guanidin D. Grupi tiol E. Grupi amino
2. Zgjidhni përgjigjet e sakta.
Aminoacidet me radikale polare të pangarkuara janë:
A. Cis B. Asn
B. Glu G. Tre
3. Zgjidhni përgjigjet e sakta.
Radikalet e aminoacideve:
A. Siguroni specifikën e strukturës parësore B. Merrni pjesë në formimin e strukturës terciare
B. Të vendosura në sipërfaqen e proteinës, ndikojnë në tretshmërinë e saj D. Formojnë qendrën aktive
D. Merrni pjesë në formimin e lidhjeve peptide
4. Zgjidhni përgjigjet e sakta.
Ndërveprimet hidrofobike mund të formohen midis radikaleve të aminoacideve:
A. Tre Lay B. Pro Tre
B. Met Ile G. Tir Ala D. Val Fen
5. Zgjidhni përgjigjet e sakta.
Lidhjet jonike mund të formohen midis radikaleve të aminoacideve:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Prill
6. Zgjidhni përgjigjet e sakta.
Lidhjet e hidrogjenit mund të formohen midis radikaleve të aminoacideve:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Ndeshje.
Lloji i lidhjes që përfshihet në formimin e strukturës së proteinave:
A. Struktura primare B. Struktura dytësore
B. Struktura terciare
D. Struktura supersekondare E. Konformimi.
1. Lidhjet hidrogjenore midis atomeve të shtyllës kurrizore peptide
2. Lidhje të dobëta ndërmjet grupeve funksionale të radikaleve të aminoacideve
3. Lidhjet ndërmjet grupeve α-amino dhe α-karboksil të aminoacideve
8. Zgjidhni përgjigjet e sakta. Tripsina:
A. Enzima proteolitike B. Përmban dy domene
B. Hidrolizon niseshtenë
D. Faqja aktive ndodhet midis domeneve. D. Përbëhet nga dy vargje polipeptide.
9. Zgjidhni përgjigjet e sakta. Atropina:
A. Neurotransmetues
B. Analog strukturor i acetilkolinës
B. Ndërvepron me receptorët H-kolinergjikë
D. Forcon përcjelljen e impulseve nervore nëpërmjet sinapseve kolinergjike
D. Frenues konkurrues i receptorëve M-kolinergjikë
10. Zgjidhni pohimet e sakta. Në proteina:
A. Struktura primare përmban informacion rreth strukturës së faqes së saj aktive
B. Qendra aktive formohet në nivel të strukturës parësore
B. Konformacioni është fiksuar në mënyrë të ngurtë nga lidhje kovalente
D. Vendi aktiv mund të ndërveprojë me një grup ligandësh të ngjashëm
për shkak të qëndrueshmërisë konformuese të proteinave D. Ndryshimet në mjedis mund të ndikojnë në afinitetin e aktivit
qendër në ligand
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
TERMAT DHE KONCEPTET THEMELORE
1. Proteina, polipeptide, aminoacide
2. Strukturat e proteinave primare, sekondare, terciare
3. Konformimi, konformimi i proteinave vendase
4. Lidhje kovalente dhe të dobëta në proteina
5. Qëndrueshmëria konformative
6. Vend aktiv i proteinave
7. Ligandët
8. Palosja e proteinave
9. Analoge strukturore te ligandeve
10. Proteinat e domenit
11. Proteinat e thjeshta dhe komplekse
12. Denatyrim i proteinave, agjentë denatyrues
13. Riaktivizimi i proteinave
Zgjidh problemet
"Organizimi strukturor i proteinave dhe baza e funksionimit të tyre"
1. Funksioni kryesor i proteinës - hemoglobina A (HbA) është transporti i oksigjenit në inde. Në popullatën njerëzore njihen forma të shumta të kësaj proteine me veti dhe funksione të ndryshuara - të ashtuquajturat hemoglobina jonormale. Për shembull, hemoglobina S, e gjetur në qelizat e kuqe të gjakut të pacientëve me sëmundje drapërocitare (HbS), është gjetur të ketë tretshmëri të ulët në kushtet e presionit të ulët të pjesshëm të oksigjenit (siç është rasti në gjakun venoz). Kjo çon në formimin e agregateve të kësaj proteine. Proteina humbet funksionin e saj, precipiton dhe qelizat e kuqe të gjakut marrin formë të çrregullt (disa prej tyre formojnë një formë drapëri) dhe shkatërrohen më shpejt se zakonisht në shpretkë. Si rezultat, zhvillohet anemia drapërocitare.
I vetmi ndryshim në strukturën primare të HbA u gjet në rajonin N-terminal të zinxhirit β të hemoglobinës. Krahasoni rajonet N-terminale të vargut β dhe tregoni se si ndryshimet në strukturën parësore të proteinës ndikojnë në vetitë dhe funksionet e saj.
 Për ta bërë këtë:
Për ta bërë këtë:
a) shkruani formulat e aminoacideve me të cilat ndryshon HbA dhe krahasoni vetitë e këtyre aminoacideve (polariteti, ngarkesa).
b) nxirret një përfundim për arsyen e uljes së tretshmërisë dhe ndërprerjes së transportit të oksigjenit në inde.
2. Figura tregon një diagram të strukturës së një proteine që ka një qendër lidhëse me një ligand (qendër aktive). Shpjegoni pse proteina është selektive në zgjedhjen e ligandit. Për ta bërë këtë:
a) mbani mend se cila është qendra aktive e një proteine dhe merrni parasysh strukturën e qendrës aktive të proteinës së treguar në figurë;
b) të shkruajë formulat e radikaleve të aminoacideve që përbëjnë qendrën aktive;
c) vizatoni një ligand që mund të ndërveprojë në mënyrë specifike me zonën aktive të proteinës. Tregoni mbi të grupet funksionale që mund të krijojnë lidhje me radikalet e aminoacideve që përbëjnë qendrën aktive;
d) tregoni llojet e lidhjeve që lindin midis ligandit dhe radikaleve aminoacide të qendrës aktive;
e) shpjegoni se në çfarë bazohet specifika e ndërveprimit protein-ligand.
 3.
Figura tregon vendin aktiv të proteinës dhe disa ligandëve.
3.
Figura tregon vendin aktiv të proteinës dhe disa ligandëve.
Përcaktoni se cili ligand ka më shumë gjasa të ndërveprojë me zonën aktive të proteinës dhe pse.
 Cilat lloje të lidhjeve lindin gjatë formimit të një kompleksi protein-ligand?
Cilat lloje të lidhjeve lindin gjatë formimit të një kompleksi protein-ligand?
4. Analogët strukturorë të ligandëve të proteinave natyrore mund të përdoren si ilaçe për të modifikuar aktivitetin e proteinave.
Acetilkolina është një ndërmjetës i transmetimit të ngacmimit në sinapset neuromuskulare. Kur acetilkolina ndërvepron me proteinat - receptorët e membranës postsinaptike të muskujve skeletorë, kanalet jonike hapen dhe ndodh tkurrja e muskujve. Ditilin është një ilaç që përdoret në disa operacione për të relaksuar muskujt, pasi prish transmetimin e impulseve nervore përmes sinapseve neuromuskulare. Shpjegoni mekanizmin e veprimit të ditilinës si relaksues i muskujve. Për ta bërë këtë:
a) shkruani formulat e acetilkolinës dhe ditilinës dhe krahasoni strukturën e tyre;
b) të përshkruajë mekanizmin e efektit relaksues të ditilinës.
5. Në disa sëmundje, temperatura e trupit të pacientit rritet, gjë që konsiderohet si një reagim mbrojtës i trupit. Megjithatë, temperaturat e larta janë të dëmshme për proteinat e trupit. Shpjegoni pse në temperatura mbi 40 °C funksioni i proteinave prishet dhe lind një kërcënim për jetën e njeriut. Për ta bërë këtë, mbani mend:
1) Struktura e proteinave dhe lidhjet që mbajnë strukturën e saj në konformacionin vendas;
2) Si ndryshon struktura dhe funksioni i proteinave me rritjen e temperaturës?;
3) Çfarë është homeostaza dhe pse është e rëndësishme për ruajtjen e shëndetit të njeriut.
Njësia modulare 2 PROTEINAT OLIGOMERIKE SI OBJEKTET E NDIKIMIT RREGULLATORË. DIVERSITETI STRUKTURAL DHE FUNKSIONAL I PROTEINAVE. METODAT PËR NDARJEN DHE PASTRIMIN E PROTEINAVE
Objektivat e mësimit Të jetë i aftë:
1. Përdor njohuritë për veçoritë e strukturës dhe funksioneve të proteinave oligomerike për të kuptuar mekanizmat adaptues të rregullimit të funksioneve të tyre.
2. Shpjegoni rolin e kaperoneve në sintezën dhe ruajtjen e konformacionit të proteinave në kushte qelizore.
3. Shpjegoni shumëllojshmërinë e manifestimeve të jetës nga shumëllojshmëria e strukturave dhe funksioneve të proteinave të sintetizuara në trup.
4. Analizoni marrëdhënien midis strukturës së proteinave dhe funksionit të tyre duke përdorur shembuj të krahasimit të hemoproteinave të lidhura - mioglobinës dhe hemoglobinës, si dhe përfaqësuesve të pesë klasave të proteinave të familjes së imunoglobulinave.
5. Zbatoni njohuritë për veçoritë e vetive fizike dhe kimike të proteinave për të zgjedhur metodat e pastrimit të tyre nga proteinat dhe papastërtitë e tjera.
6. Interpretoni rezultatet e përbërjes sasiore dhe cilësore të proteinave të plazmës së gjakut për të konfirmuar ose sqaruar diagnozën klinike.
Dije:
1. Veçoritë e strukturës së proteinave oligomerike dhe mekanizmat adaptues për rregullimin e funksioneve të tyre duke përdorur shembullin e hemoglobinës.
2. Struktura dhe funksionet e kaperoneve dhe rëndësia e tyre për ruajtjen e konformacionit vendas të proteinave në kushte qelizore.
3. Parimet e kombinimit të proteinave në familje bazuar në ngjashmërinë e konformimit dhe funksioneve të tyre duke përdorur shembullin e imunoglobulinave.
4. Metodat për ndarjen e proteinave bazuar në karakteristikat e vetive fiziko-kimike të tyre.
5. Elektroforeza e plazmës së gjakut si metodë për vlerësimin e përbërjes cilësore dhe sasiore të proteinave.
TEMA 1.4. TIPARET E STRUKTURËS DHE FUNKSIONIMIT TË PROTEINAVE OLIGOMERIKE DUKE PËRDORUR SHEMBULLIN E HEMOGLOBINËS
1. Shumë proteina përmbajnë disa zinxhirë polipeptidikë. Proteinat e tilla quhen oligomerike, dhe zinxhirë individualë - protomerët. Protomerët në proteinat oligomerike lidhen me shumë lidhje të dobëta jokovalente (hidrofobike, jonike, hidrogjen). Ndërveprim
protomerët kryhen në sajë të komplementariteti sipërfaqet kontaktuese të tyre.
Numri i protomerëve në proteinat oligomerike mund të ndryshojë shumë: hemoglobina përmban 4 protomere, enzima aspartate aminotransferaza ka 12 protomere dhe proteina e virusit të mozaikut të duhanit përmban 2120 protomere të lidhur me lidhje jokovalente. Rrjedhimisht, proteinat oligomerike mund të kenë pesha shumë të larta molekulare.
Ndërveprimi i një protomeri me të tjerët mund të konsiderohet një rast i veçantë i ndërveprimit protein-ligand, pasi secili protomer shërben si ligand për protomerët e tjerë. Numri dhe mënyra e bashkimit të protomerëve në një proteinë quhet struktura e proteinave kuaternare.
Proteinat mund të përmbajnë protomerë të strukturave të njëjta ose të ndryshme, për shembull, homodimerët janë proteina që përmbajnë dy protomerë identikë, dhe heterodimerët janë proteina që përmbajnë dy protomerë të ndryshëm.
Nëse proteinat përmbajnë protomerë të ndryshëm, atëherë mbi to mund të formohen qendra lidhëse me ligandë të ndryshëm që ndryshojnë në strukturë. Kur një ligand lidhet me zonën aktive, funksioni i kësaj proteine manifestohet. Një qendër e vendosur në një protomer të ndryshëm quhet alosterike (e ndryshme nga ajo aktive). Duke kontaktuar ligand ose efektor alosterik, ai kryen një funksion rregullues (Fig. 1.18). Ndërveprimi i qendrës alosterike me efektorin shkakton ndryshime konformacionale në strukturën e të gjithë proteinës oligomerike për shkak të qëndrueshmërisë së saj konformuese. Kjo ndikon në afinitetin e zonës aktive për një ligand specifik dhe rregullon funksionin e asaj proteine. Një ndryshim në konformacionin dhe funksionin e të gjithë protomerëve gjatë ndërveprimit të një proteine oligomerike me të paktën një ligand quhet ndryshime konformative bashkëpunuese. Efektorët që përmirësojnë funksionin e proteinave quhen aktivizues, dhe efektorët që pengojnë funksionin e tij janë
frenuesit.
 Kështu, proteinat oligomerike, si dhe proteinat me strukturë domeni, kanë një veti të re në krahasim me proteinat monomerike - aftësinë për të rregulluar në mënyrë alosterike funksionet (rregullimi duke i bashkangjitur ligandëve të ndryshëm proteinës). Kjo mund të shihet duke krahasuar strukturat dhe funksionet e dy proteinave komplekse të lidhura ngushtë, mioglobinës dhe hemoglobinës.
Kështu, proteinat oligomerike, si dhe proteinat me strukturë domeni, kanë një veti të re në krahasim me proteinat monomerike - aftësinë për të rregulluar në mënyrë alosterike funksionet (rregullimi duke i bashkangjitur ligandëve të ndryshëm proteinës). Kjo mund të shihet duke krahasuar strukturat dhe funksionet e dy proteinave komplekse të lidhura ngushtë, mioglobinës dhe hemoglobinës.
Oriz. 1.18. Skema e strukturës së një proteine dimerike
2. Formimi i strukturave hapësinore dhe funksionimi i mioglobinës.
Mioglobina (Mb) është një proteinë që gjendet në muskujt e kuq, funksioni kryesor i së cilës është krijimi i rezervave O 2 të nevojshme për punën intensive të muskujve. Mb është një proteinë komplekse që përmban një pjesë proteinike - apoMb dhe një pjesë jo proteinike - hem. Struktura primare e apoMB përcakton konformacionin e saj kompakt globular dhe strukturën e qendrës aktive, në të cilën është ngjitur pjesa joproteinike e mioglobinës, hemi. Oksigjeni që vjen nga gjaku në muskuj lidhet me hemet Fe+2 në mioglobinë. Mb është një proteinë monomerike që ka një afinitet shumë të lartë për O 2, prandaj lirimi i oksigjenit nga mioglobina ndodh vetëm gjatë punës intensive muskulare, kur presioni i pjesshëm i O 2 zvogëlohet ndjeshëm. Në muskujt e kuq, në ribozomet gjatë përkthimit, sintetizohet struktura primare e MB, e përfaqësuar nga një sekuencë specifike prej 153 mbetjesh aminoacide. Struktura dytësore e Mb përmban tetë α-spira, të quajtura me shkronja latine nga A në H, midis të cilave ka rajone jo-spirale. Struktura terciare e Mb ka formën e një rruzulli të ngjeshur, në vrimën e së cilës ndodhet qendra aktive midis helicave F dhe E (Fig. 1.19).
 Oriz. 1.19. Struktura e mioglobinës
Oriz. 1.19. Struktura e mioglobinës
3. Veçoritë e strukturës dhe funksionimit të qendrës aktive të TM. Qendra aktive e Mb formohet kryesisht nga radikalet hidrofobike të aminoacideve, të vendosura gjerësisht nga njëri-tjetri në strukturën parësore (për shembull, Tri 3 9 dhe Fen 138) Ligandët e dobët të tretshëm në ujë - heme dhe O 2 - ngjiten në qendrën aktive. Hemi është një ligand specifik i apoMB (Fig. 1.20), baza e të cilit përbëhet nga katër unaza piroli të lidhura me ura metenil; në qendër është një atom Fe+ 2 i lidhur me atomet e azotit të unazave të pirrolit me katër lidhje koordinimi. Në qendrën aktive të Mb, përveç radikaleve të aminoacideve hidrofobike, ka edhe mbetje të dy aminoacideve me radikale hidrofile - Gis E 7(Gis 64) dhe
 GIS F 8
GIS F 8
 (93 e tij) (Fig. 1.21).
(93 e tij) (Fig. 1.21).
Oriz. 1.20. Struktura e hemit - pjesa jo proteinike e mioglobinës dhe hemoglobinës
Gis E 7 Oriz. 1.21. Vendndodhja e hemit dhe O2 në vendin aktiv të apomioglobinës dhe protomerëve të hemoglobinës Hemi është një ligand specifik i apoMB (Fig. 1.20), baza e të cilit përbëhet nga katër unaza piroli të lidhura me ura metenil; Hemi është i lidhur në mënyrë kovalente me F8 të tij nëpërmjet një atomi hekuri. O 2 ngjitet në hekurin në anën tjetër të planit të hemit. E 7 e tij është e nevojshme për orientimin e saktë të O 2 dhe lehtëson shtimin e oksigjenit në hemin Fe + 2
formon një lidhje koordinimi me Fe+ 2 dhe fikson fort hemin në qendrën aktive.
e nevojshme për orientimin e saktë në qendrën aktive të një ligandi tjetër - O 2 gjatë ndërveprimit të tij me hemin Fe + 2. Mikromjedisi i hemit krijon kushte për lidhje të fortë, por të kthyeshme të O 2 me Fe + 2 dhe parandalon që uji të hyjë në zonën aktive hidrofobike, gjë që mund të çojë në oksidimin e tij në Fe + 3.- një familje proteinash, si mioglobina, e lidhur me proteinat komplekse (hemoproteinat). Ata kanë një strukturë tetramerike dhe përmbajnë dy zinxhirë α, por ndryshojnë në strukturën e dy zinxhirëve të tjerë polipeptidikë (zinxhirët 2α-, 2x). Struktura e zinxhirit të dytë polipeptid përcakton veçoritë e funksionimit të këtyre formave të Hb. Rreth 98% e hemoglobinës në qelizat e kuqe të gjakut të një të rrituri është hemoglobina A(2α-, 2p-zinxhirë).
Gjatë zhvillimit të fetusit, funksionojnë dy lloje kryesore të hemoglobinës: Hb embrional(2α, 2ε), e cila gjendet në fazat e hershme të zhvillimit të fetusit, dhe hemoglobina F (fetale)- (2α, 2γ), e cila zëvendëson hemoglobinën e hershme fetale në muajin e gjashtë të zhvillimit intrauterin dhe vetëm pas lindjes zëvendësohet nga Hb A.
Hb A është një proteinë e lidhur me mioglobinën (Mb) që gjendet në qelizat e kuqe të gjakut të rritur. Struktura e protomerëve të tij individualë është e ngjashme me atë të mioglobinës. Strukturat dytësore dhe terciare të protomerëve të mioglobinës dhe hemoglobinës janë shumë të ngjashme, pavarësisht nga fakti se në strukturën parësore të zinxhirëve të tyre polipeptidë vetëm 24 mbetje aminoacide janë identike (struktura dytësore e protomerëve të hemoglobinës, si mioglobina, përmban tetë α-helika, caktohet me shkronja latine nga A në H, dhe struktura terciare ka formën e një rruzulli kompakt). Por ndryshe nga mioglobina, hemoglobina ka një strukturë oligomerike, e përbërë nga katër zinxhirë polipeptidikë të lidhur me lidhje jo kovalente (Figura 1.22).
Çdo protomer Hb shoqërohet me një pjesë jo proteinike - hemin dhe protomerët fqinjë. Lidhja e pjesës proteinike të Hb me hemin është e ngjashme me atë të mioglobinës: në qendrën aktive të proteinës, pjesët hidrofobike të hemit janë të rrethuara nga radikale aminoacide hidrofobike me përjashtim të His F 8 dhe His E 7, të cilat ndodhen në të dy anët e rrafshit të hemit dhe luajnë një rol të ngjashëm në funksionimin e proteinës dhe lidhjen e saj me oksigjenin (shih strukturën e mioglobinës).
 Oriz. 1.22. Struktura oligomerike e hemoglobinës
Oriz. 1.22. Struktura oligomerike e hemoglobinës
Përveç kësaj, Hemi është një ligand specifik i apoMB (Fig. 1.20), baza e të cilit përbëhet nga katër unaza piroli të lidhura me ura metenil; kryen një të rëndësishme rol shtesë në funksionimin e Nv. Hemi i lirë ka një afinitet 25,000 herë më të lartë për CO se sa për O2.
5. Funksioni kryesor i HB është transporti i O2 nga mushkëritë në inde. Ndryshe nga mioglobina monomerike, e cila ka një afinitet shumë të lartë për O2 dhe kryen funksionin e ruajtjes së oksigjenit në muskujt e kuq, struktura oligomerike e hemoglobinës siguron:
1) ngopja e shpejtë e HB me oksigjen në mushkëri;
2) aftësia e HB për të lëshuar oksigjen në inde me një presion të pjesshëm relativisht të lartë prej O 2 (20-40 mm Hg);
3) mundësia e rregullimit të afinitetit të Hb ndaj O2.
6. Ndryshimet bashkëpunuese në konformacionin e protomerëve të hemoglobinës përshpejtojnë lidhjen e O 2 në mushkëri dhe çlirimin e tij në inde. Në mushkëri, presioni i lartë i pjesshëm i O2 nxit lidhjen e tij me Hb në vendin aktiv të katër protomerëve (2α dhe 2β). Qendra aktive e çdo protomeri, si në mioglobinë, ndodhet midis dy α-helikave (F dhe E) në një xhep hidrofobik. Ai përmban një pjesë jo proteinike - hemin, të lidhur me pjesën proteinike nga shumë ndërveprime të dobëta hidrofobike dhe një lidhje të fortë midis Fe 2 + hemit dhe His F 8 (shih Fig. 1.21).
Në deoksihemoglobinë, për shkak të kësaj lidhjeje me His F 8, atomi Fe 2 + del nga rrafshi i hemit drejt histidinës. Lidhja e O 2 me Fe 2 + ndodh në anën tjetër të hemit në rajonin His E 7 duke përdorur një lidhje të vetme koordinimi të lirë. E 7 e tij siguron kushte optimale për lidhjen e O 2 me hekurin hem.
Shtimi i O 2 në atomin Fe + 2 të një protomeri shkakton lëvizjen e tij në rrafshin e hemit, e ndjekur nga mbetjet e histidinës që lidhen me të.
 Oriz. 1.23. Ndryshimi në konformacionin e protomerit të hemoglobinës kur kombinohet me O2
Oriz. 1.23. Ndryshimi në konformacionin e protomerit të hemoglobinës kur kombinohet me O2
Kjo çon në një ndryshim në konformimin e të gjithë zinxhirëve polipeptidikë për shkak të qëndrueshmërisë së tyre konformuese. Ndryshimi i konformacionit të zinxhirëve të tjerë lehtëson ndërveprimin e tyre me molekulat e mëvonshme O 2.
Molekula e katërt O 2 ngjitet në hemoglobinë 300 herë më lehtë se e para (Fig. 1.24).
 Oriz. 1.24. Ndryshimet bashkëpunuese në konformimin e protomerëve të hemoglobinës gjatë ndërveprimit të saj me O2
Oriz. 1.24. Ndryshimet bashkëpunuese në konformimin e protomerëve të hemoglobinës gjatë ndërveprimit të saj me O2
Në inde, çdo molekulë e mëvonshme O 2 shkëputet më lehtë se ajo e mëparshme, gjithashtu për shkak të ndryshimeve bashkëpunuese në konformacionin e protomerëve.
7. CO 2 dhe H+, të formuara gjatë katabolizmit të substancave organike, zvogëlojnë afinitetin e hemoglobinës për O 2 në raport me përqendrimin e tyre. Energjia e nevojshme për funksionimin e qelizave prodhohet kryesisht në mitokondri gjatë oksidimit të substancave organike duke përdorur O 2 të lëshuar nga mushkëritë nga hemoglobina. Si rezultat i oksidimit të substancave organike, formohen produktet përfundimtare të zbërthimit të tyre: CO 2 dhe K 2 O, sasia e të cilave është në përpjesëtim me intensitetin e proceseve të oksidimit në vazhdim.
CO 2 shpërndahet nga qelizat në gjak dhe depërton në qelizat e kuqe të gjakut, ku, nën veprimin e enzimës karbanhidrazë, shndërrohet në acid karbonik. Ky acid i dobët shpërndahet në një proton dhe një jon bikarbonat.
H+ janë të aftë të bashkohen me radikalët e Tij 14 6 në zinxhirët α- dhe β të hemoglobinës, d.m.th. në zonat larg hemit. Protonizimi i hemoglobinës redukton afinitetin e saj për O 2, nxit largimin e O 2 nga oxyHb, formimin e deoksiHb dhe rrit furnizimin me oksigjen në inde në raport me numrin e protoneve të formuara (Fig. 1.25).
Një rritje në sasinë e oksigjenit të çliruar në varësi të rritjes së përqendrimit të H+ në qelizat e kuqe të gjakut quhet efekti Bohr (emërtuar sipas fiziologut danez Christian Bohr, i cili zbuloi i pari këtë efekt).
Në mushkëri, një presion i lartë i pjesshëm i oksigjenit nxit lidhjen e tij me deoksiHb, gjë që redukton afinitetin e proteinës për H +. Protonet e liruara nën veprimin e acidit karbonik reagojnë me bikarbonatet për të formuar CO 2 dhe H 2 O

 Oriz. 1.25. Varësia e afinitetit të Hb për O 2 nga përqendrimi i CO 2 dhe protoneve (efekti Bohr):
Oriz. 1.25. Varësia e afinitetit të Hb për O 2 nga përqendrimi i CO 2 dhe protoneve (efekti Bohr):
A- ndikimi i përqendrimit të CO 2 dhe H+ në çlirimin e O 2 nga kompleksi me HB (efekti Bohr); B- oksigjenimi i deoksihemoglobinës në mushkëri, formimi dhe çlirimi i CO 2.
CO 2 që rezulton hyn në hapësirën alveolare dhe hiqet me ajër të nxjerrë. Kështu, sasia e oksigjenit të çliruar nga hemoglobina në inde rregullohet nga produktet e katabolizmit të substancave organike: sa më intensiv të jetë shpërbërja e substancave, për shembull gjatë ushtrimeve fizike, aq më i lartë është përqendrimi i CO 2 dhe H + dhe aq më shumë oksigjen. indet marrin si rezultat i uljes së afinitetit të Hb ndaj O2.
8. Rregullimi alosterik i afinitetit të Hb për O2 nga ligandi - 2,3-bisfosfoglicerati. Në eritrocite, ligandi alosterik i hemoglobinës, 2,3-bisfosfoglicerati (2,3-BPG), sintetizohet nga produkti i oksidimit të glukozës - 1,3-bisfosfoglicerati. Në kushte normale, përqendrimi i 2,3-BPG është i lartë dhe i krahasueshëm me përqendrimin e Hb. 2,3-BPG ka një ngarkesë të fortë negative prej -5.
 Bisfosfoglicerati në kapilarët e indeve, duke u lidhur me deoksihemoglobinën, rrit çlirimin e oksigjenit në inde, duke reduktuar afinitetin e Hb për O 2.
Bisfosfoglicerati në kapilarët e indeve, duke u lidhur me deoksihemoglobinën, rrit çlirimin e oksigjenit në inde, duke reduktuar afinitetin e Hb për O 2.
Në qendër të molekulës tetramerike të hemoglobinës është një zgavër. Ai formohet nga mbetjet aminoacide të të katër protomerëve (shih Fig. 1.22). Në kapilarët e indeve, protonizimi i Hb (efekti Bohr) çon në këputjen e lidhjes midis hekurit heme dhe O2. Në një molekulë
deoksihemoglobina, në krahasim me oksihemoglobinën, shfaqen lidhje shtesë jonike që lidhin protomerët, si rezultat i të cilave përmasat e zgavrës qendrore rriten në krahasim me oksihemoglobinën. Kaviteti qendror është vendi i lidhjes së 2,3-BPG me hemoglobinën. Për shkak të ndryshimit në madhësinë e zgavrës qendrore, 2,3-BPG mund të lidhet vetëm me deoksihemoglobinën.
2,3-BPG ndërvepron me hemoglobinën në një vend të largët nga qendrat aktive të proteinës dhe i përket alosterike ligandët (rregullatorë), dhe kaviteti qendror i Hb është qendër alosterike. 2,3-BPG ka një ngarkesë të fortë negative dhe ndërvepron me pesë grupe të ngarkuara pozitivisht të dy zinxhirëve β të Hb: grupi α-amino terminal N-të Val dhe radikalët Lys 82 His 143 (Fig. 1.26).
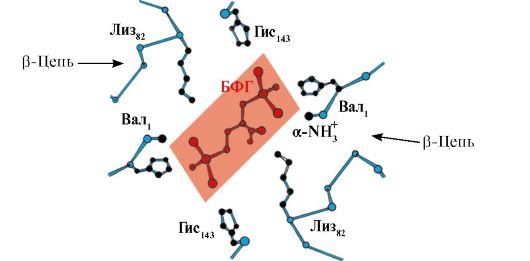 Oriz. 1.26. BPG në zgavrën qendrore të deoksihemoglobinës
Oriz. 1.26. BPG në zgavrën qendrore të deoksihemoglobinës
BPG lidhet me tre grupe të ngarkuara pozitivisht në secilën β-fillesë.
Në kapilarët e indeve, deoksihemoglobina që rezulton ndërvepron me 2,3-BPG dhe formohen lidhje jonike midis radikalëve të ngarkuar pozitivisht të zinxhirëve β dhe ligandit të ngarkuar negativisht, të cilët ndryshojnë konformimin e proteinës dhe zvogëlojnë afinitetin e Hb për O. 2 . Një rënie në afinitetin e Hb për O 2 nxit një çlirim më efikas të O 2 në inde.
Në mushkëri, në presion të lartë të pjesshëm, oksigjeni ndërvepron me Hb, duke bashkuar hekurin hem; në këtë rast, konformacioni i proteinës ndryshon, zgavra qendrore zvogëlohet dhe 2,3-BPG zhvendoset nga qendra alosterike.
Kështu, proteinat oligomerike kanë veti të reja në krahasim me proteinat monomerike. Ngjitja e ligandëve në vende
larg nga njëra-tjetra në hapësirë (allosterike), mund të shkaktojnë ndryshime konformacionale në të gjithë molekulën e proteinës. Për shkak të ndërveprimit me ligandët rregullator, ndodh një ndryshim në konformimin dhe përshtatja e funksionit të molekulës së proteinës ndaj ndryshimeve mjedisore.
TEMA 1.5. MIRËMBAJTJA E KONFORMACIONIT VENDOR TË PROTEINAVE NË KUSHTET QELIZORE
Në qeliza, gjatë sintezës së vargjeve polipeptide, transporti i tyre përmes membranave në pjesët përkatëse të qelizës, gjatë procesit të palosjes (formimit të konformacionit vendas) dhe gjatë montimit të proteinave oligomerike, si dhe gjatë funksionimit të tyre, ndërmjetësohet. , në strukturën e proteinave lindin konformacione të prirura për grumbullim, të paqëndrueshme. Radikalet hidrofobike, zakonisht të fshehura brenda molekulës së proteinës në konformacionin vendas, shfaqen në sipërfaqe në një konformacion të paqëndrueshëm dhe priren të kombinohen me grupe proteinash të tjera që janë pak të tretshme në ujë. Në qelizat e të gjithë organizmave të njohur janë gjetur proteina të veçanta që sigurojnë palosjen optimale të proteinave qelizore, stabilizojnë konformimin e tyre vendas gjatë funksionimit dhe, më e rëndësishmja, ruajnë strukturën dhe funksionet e proteinave ndërqelizore kur homeostaza është e shqetësuar. Këto proteina quhen "kaperone" që do të thotë "dado" në frëngjisht.
1. Kaperonët molekularë dhe roli i tyre në parandalimin e denatyrimit të proteinave.
Kaperonët (CH) klasifikohen sipas masës së nënnjësive të tyre. Kaperonët me peshë të lartë molekulare kanë një masë nga 60 deri në 110 kDa. Ndër to, tre klasa janë më të studiuara: Sh-60, Sh-70 dhe Sh-90. Çdo klasë përfshin një familje proteinash të lidhura. Kështu, Sh-70 përmban proteina me një peshë molekulare nga 66 në 78 kDa. Kaperonët me peshë të ulët molekulare kanë një peshë molekulare nga 40 deri në 15 kDa.
Midis shoqëruesve ka konstituive proteinat, sinteza e lartë bazale e të cilave nuk varet nga efektet e stresit në qelizat e trupit, dhe e induktueshme, sinteza e të cilave në kushte normale është e dobët, por rritet ndjeshëm nën stres. Chaperonet e induktueshme quhen gjithashtu "proteina të goditjes nga nxehtësia" sepse ato u zbuluan fillimisht në qelizat e ekspozuara ndaj temperaturave të larta. Në qeliza, për shkak të përqendrimit të lartë të proteinave, riaktivizimi spontan i proteinave pjesërisht të denatyruara është i vështirë. Sh-70 mund të parandalojë fillimin e denatyrimit dhe të ndihmojë në rikthimin e konformacionit vendas të proteinave. Kaperone molekulare-70- një klasë shumë e konservuar proteinash që gjenden në të gjitha pjesët e qelizës: citoplazmë, bërthamë, rrjetë endoplazmatike, mitokondri. Në skajin karboksil të zinxhirit të vetëm polipeptid Ш-70 ekziston një zonë që është një brazdë e aftë të ndërveprojë me peptidet me gjatësi
nga 7 deri në 9 mbetje aminoacide të pasuruara me radikale hidrofobike. Rajone të tilla në proteinat globulare ndodhin afërsisht çdo 16 aminoacide. Sh-70 është në gjendje të mbrojë proteinat nga inaktivizimi i temperaturës dhe të rivendosë konformimin dhe aktivitetin e proteinave pjesërisht të denatyruara.
2. Roli i kaperonëve në palosjen e proteinave. Gjatë sintezës së proteinave në ribozomë, rajoni N-terminal i polipeptidit sintetizohet para atij C-terminal. Për të formuar konformacionin vendas, kërkohet sekuenca e plotë e aminoacideve të proteinës. Në procesin e sintezës së proteinave, kaperonet-70, për shkak të strukturës së qendrës së tyre aktive, janë në gjendje të mbyllin zonat e polipeptidit që janë të prirura për grumbullim, të pasuruara me radikale aminoacide hidrofobike derisa të përfundojë sinteza (Figura 1.27, A. ).
 Oriz. 1.27. Pjesëmarrja e kaperonëve në palosjen e proteinave
Oriz. 1.27. Pjesëmarrja e kaperonëve në palosjen e proteinave
A - pjesëmarrja e chaperones-70 në parandalimin e ndërveprimeve hidrofobike midis seksioneve të polipeptidit të sintetizuar; B - formimi i konformacionit vendas të proteinës në kompleksin chaperone
Shumë proteina me molekulare të lartë që kanë një konformacion kompleks, siç është një strukturë domeni, palosen në një hapësirë të veçantë të formuar nga Sh-60. Ш-60 funksionon si një kompleks oligomerik i përbërë nga 14 nënnjësi. Ata formojnë dy unaza të zbrazëta, secila prej të cilave përbëhet nga shtatë nënnjësi, këto unaza janë të lidhura me njëra-tjetrën. Çdo nënnjësi Sh-60 përbëhet nga tre fusha: apikale (apikale), e pasuruar me radikale hidrofobike përballë zgavrës së unazës, e ndërmjetme dhe ekuatoriale (Fig. 1.28).
 Oriz. 1.28. Struktura e kompleksit chaperonin e përbërë nga 14 Ш-60
Oriz. 1.28. Struktura e kompleksit chaperonin e përbërë nga 14 Ш-60
A - pamje anësore; B - pamje nga lart
Proteinat e sintetizuara, të cilat kanë elementë në sipërfaqe, karakteristikë të molekulave të shpalosura, në veçanti radikalet hidrofobike, hyjnë në zgavrën e unazave të pasagjerëve. Në mjedisin specifik të këtyre zgavrave numërohen konformacionet e mundshme derisa të gjendet e vetmja që është energjikisht më e favorshme (Fig. 1.27, B). Formimi i konformacioneve dhe çlirimi i proteinave shoqërohet me hidrolizë të ATP në rajonin ekuatorial. Në mënyrë tipike, një palosje e tillë e varur nga personeli kërkon një sasi të konsiderueshme energjie.
Përveç pjesëmarrjes në formimin e strukturës tredimensionale të proteinave dhe ripërtëritjen e proteinave pjesërisht të denatyruara, kaperonët janë gjithashtu të nevojshëm për shfaqjen e proceseve të tilla themelore si grumbullimi i proteinave oligomerike, njohja dhe transportimi i proteinave të denatyruara në lizozome. transporti i proteinave nëpër membrana dhe pjesëmarrja në rregullimin e aktivitetit të komplekseve proteinike.
TEMA 1.6. NDRYSHIM I PROTEINAVE. FAMILJET PROTEINARE: SHEMBULL I IMUNOGLOBULINAVE
1. Proteinat luajnë një rol vendimtar në jetën e qelizave individuale dhe të gjithë organizmit shumëqelizor, dhe funksionet e tyre janë çuditërisht të ndryshme. Kjo përcaktohet nga karakteristikat e strukturës primare dhe konformacioneve të proteinave, struktura unike e qendrës aktive dhe aftësia për të lidhur ligandët specifikë.
Vetëm një pjesë shumë e vogël e të gjitha varianteve të mundshme të zinxhirëve peptidë mund të adoptojnë një strukturë hapësinore të qëndrueshme; shumicë
prej tyre mund të marrin shumë konformacione me përafërsisht të njëjtën energji Gibbs, por me veti të ndryshme. Struktura primare e proteinave më të njohura të përzgjedhura nga evolucioni biologjik siguron një stabilitet të jashtëzakonshëm të një prej konformacioneve, i cili përcakton karakteristikat e funksionimit të kësaj proteine.
2. Familjet proteinike. Brenda të njëjtës specie biologjike, zëvendësimet e mbetjeve të aminoacideve mund të çojnë në shfaqjen e proteinave të ndryshme që kryejnë funksione të lidhura dhe kanë sekuenca homologe të aminoacideve. Proteina të tilla të lidhura kanë konformacione jashtëzakonisht të ngjashme: numri dhe pozicioni relativ i α-helikave dhe/ose strukturave β, shumica e kthesave dhe kthesave të zinxhirëve polipeptidikë janë të ngjashëm ose identikë. Proteinat me rajone homologe të zinxhirit polipeptid, konformacion të ngjashëm dhe funksione të ngjashme klasifikohen në familje proteinike. Shembuj të familjeve të proteinave: proteinazat e serinës, familja e imunoglobulinave, familja e mioglobinës.
Proteinazat serine- një familje proteinash që kryejnë funksionin e enzimave proteolitike. Këto përfshijnë enzimat tretëse - kimotripsinën, tripsinën, elastazën dhe shumë faktorë të koagulimit të gjakut. Këto proteina kanë aminoacide identike në 40% të pozicioneve të tyre dhe një konformacion shumë të ngjashëm (Fig. 1.29).

Oriz. 1.29. Strukturat hapësinore të elastazës (A) dhe kimotripsinës (B)
Disa zëvendësime të aminoacideve kanë çuar në ndryshime në specifikën e substratit të këtyre proteinave dhe në shfaqjen e diversitetit funksional brenda familjes.
3. Familja e imunoglobulinave. Proteinat e superfamiljes së imunoglobulinave, e cila përfshin tre familje proteinash, luajnë një rol të madh në funksionimin e sistemit imunitar:
Antitrupat (imunoglobulinat);
Receptorët e limfociteve T;
Proteinat e kompleksit kryesor të histokompatibilitetit - MHC klasa 1 dhe 2 (Major Histocompatibility Complex).
Të gjitha këto proteina kanë një strukturë domeni, përbëhen nga domene homologe të ngjashme me imunitetin dhe kryejnë funksione të ngjashme: ato ndërveprojnë me struktura të huaja, ose të tretura në gjak, limfë ose lëng ndërqelizor (antitrupa), ose të vendosura në sipërfaqen e qelizave (veta ose të huaj).
4. Antitrupat- proteinat specifike të prodhuara nga limfocitet B në përgjigje të hyrjes së një strukture të huaj në trup, të quajtura antigjen.
Karakteristikat e strukturës së antitrupave
Molekulat më të thjeshta të antitrupave përbëhen nga katër zinxhirë polipeptidikë: dy të lehta identike - L, që përmbajnë rreth 220 aminoacide dhe dy të rënda identike - H, të përbërë nga 440-700 aminoacide. Të katër zinxhirët në molekulën e antitrupave janë të lidhur me shumë lidhje jokovalente dhe katër lidhje disulfide (Fig. 1.30).
Zinxhirët e lehtë të antitrupave përbëhen nga dy fusha: një domen i ndryshueshëm (VL), i vendosur në rajonin N-terminal të zinxhirit polipeptid dhe një domen konstant (CL), i vendosur në fundin C. Zinxhirët e rëndë zakonisht kanë katër domene: një variabël (VH), e vendosur në skajin N dhe tre domene konstante (CH1, CH2, CH3) (shih Fig. 1.30). Çdo domen imunoglobulinash ka një superstrukturë të fletës β në të cilën dy mbetje cisteine janë të lidhura me një lidhje disulfide.
Midis dy domeneve konstante CH1 dhe CH2 ekziston një rajon që përmban një numër të madh mbetjesh proline, të cilat parandalojnë formimin e një strukture dytësore dhe ndërveprimin e zinxhirëve H fqinjë në këtë segment. Ky rajon i varur i jep molekulës së antitrupave fleksibilitet. Midis domeneve të ndryshueshme të zinxhirit të rëndë dhe të lehtë ekzistojnë dy vende identike të lidhjes së antigjenit (vendet aktive për lidhjen e antigjeneve), prandaj antitrupa të tillë shpesh quhen dyvalente. Jo e gjithë sekuenca e aminoacideve të rajoneve të ndryshueshme të të dy zinxhirëve është e përfshirë në lidhjen e antigjenit me antitrupin, por vetëm 20-30 aminoacide të vendosura në rajonet hipervariabile të secilit zinxhir. Janë këto rajone që përcaktojnë aftësinë unike të secilit lloj antitrupi për të bashkëvepruar me antigjenin plotësues përkatës.
Antitrupat janë një nga linjat e mbrojtjes së trupit kundër organizmave të huaj pushtues. Funksionimi i tyre mund të ndahet në dy faza: faza e parë është njohja dhe lidhja e antigjenit në sipërfaqen e organizmave të huaj, gjë që është e mundur për shkak të pranisë së vendeve të lidhjes së antigjenit në strukturën e antitrupave; faza e dytë është fillimi i procesit të inaktivizimit dhe shkatërrimit të antigjenit. Specifikimi i fazës së dytë varet nga klasa e antitrupave. Ekzistojnë pesë klasa të zinxhirëve të rëndë që ndryshojnë nga njëri-tjetri në strukturën e domeneve konstante: α, δ, ε, γ dhe μ, sipas të cilave dallohen pesë klasa të imunoglobulinave: A, D, E, G dhe M.
Karakteristikat strukturore të zinxhirëve të rëndë u japin rajoneve të menteshës dhe rajoneve C-terminale të zinxhirëve të rëndë një karakteristikë konformimi për secilën klasë. Pasi antigjeni lidhet me një antitrup, ndryshimet konformative në domenet konstante përcaktojnë rrugën për heqjen e antigjenit.
 Oriz. 1. 30. Struktura e domenit të IgG
Oriz. 1. 30. Struktura e domenit të IgG
Imunoglobulinat M
Imunoglobulinat M kanë dy forma.
Forma monomerike- Klasa e parë e antitrupave të prodhuar nga zhvillimi i limfociteve B. Më pas, shumë qeliza B kalojnë në prodhimin e klasave të tjera të antitrupave, por me të njëjtin vend lidhës antigjen. IgM është ngulitur në membranë dhe vepron si një receptor i njohjes së antigjenit. Integrimi i IgM në membranën qelizore është i mundur për shkak të pranisë së 25 mbetjeve të aminoacideve hidrofobike në pjesën e bishtit të rajonit.
Forma sekretore e IgM përmban pesë nënnjësi monomere të lidhura me njëra-tjetrën me lidhje disulfide dhe një zinxhir shtesë polipeptid J (Fig. 1.31). Zinxhirët e rëndë të monomerëve të kësaj forme nuk përmbajnë një bisht hidrofobik. Pentameri ka 10 vende të lidhjes së antigjenit dhe për këtë arsye është efektiv në njohjen dhe largimin e antigjenit që hyn fillimisht në trup. Forma sekretore e IgM është klasa kryesore e antitrupave të sekretuara në gjak gjatë përgjigjes imune parësore. Lidhja e IgM me një antigjen ndryshon konformacionin e IgM dhe indukton lidhjen e tij me përbërësin e parë proteinik të sistemit të komplementit (sistemi i komplementit është një grup proteinash të përfshira në shkatërrimin e antigjenit) dhe aktivizimin e këtij sistemi. Nëse antigjeni ndodhet në sipërfaqen e një mikroorganizmi, sistemi i komplementit shkakton prishje të integritetit të membranës qelizore dhe vdekjen e qelizës bakteriale.
Imunoglobulinat G
Në mënyrë sasiore, kjo klasë imunoglobulinash mbizotëron në gjak (75% e të gjitha Ig-ve). IgG - monomeret, klasa kryesore e antitrupave të sekretuara në gjak gjatë një përgjigje dytësore imune. Pasi IgG ndërvepron me antigjenet sipërfaqësore të mikroorganizmave, kompleksi antigjen-antitrup është në gjendje të lidhë dhe aktivizojë proteinat e sistemit të komplementit ose mund të ndërveprojë me receptorët specifikë të makrofagëve dhe neutrofileve. Ndërveprimi me fagocitet çon
 Oriz. 1.31. Struktura e formës sekretore të IgM
Oriz. 1.31. Struktura e formës sekretore të IgM
në përthithjen e komplekseve antigjen-antitrup dhe shkatërrimin e tyre në fagozomet qelizore. IgG është e vetmja klasë e antitrupave që janë në gjendje të depërtojnë në barrierën placentare dhe të ofrojnë mbrojtje intrauterine të fetusit nga infeksionet.
Imunoglobulinat A
Klasa kryesore e antitrupave të pranishëm në sekrecione (qumësht, pështymë, sekrecione të traktit respirator dhe traktit të zorrëve). IgA sekretohet kryesisht në formë dimerike, ku monomerët janë të lidhur me njëri-tjetrin përmes një zinxhiri J shtesë (Fig. 1.32).
IgA nuk ndërvepron me sistemin e komplementit dhe qelizat fagocitare, por duke u lidhur me mikroorganizmat, antitrupat parandalojnë lidhjen e tyre me qelizat epiteliale dhe depërtimin në trup.
Imunoglobulinat E
Imunoglobulinat E përfaqësohen nga monomere në të cilët zinxhirët e rëndë ε përmbajnë, si zinxhirët μ e imunoglobulinave M, një variabël dhe katër domene konstante. IgE pas sekretimit lidhet me të
 Oriz. 1.32. Struktura e IgA
Oriz. 1.32. Struktura e IgA
Rajonet C-terminale me receptorët përkatës në sipërfaqen e mastociteve dhe bazofileve. Si rezultat, ato bëhen receptorë për antigjenet në sipërfaqen e këtyre qelizave (Fig. 1.33).
 Oriz. 1.33. Ndërveprimi i IgE me antigjenin në sipërfaqen e një qelize mast
Oriz. 1.33. Ndërveprimi i IgE me antigjenin në sipërfaqen e një qelize mast
Pasi antigjeni ngjitet në vendet përkatëse të lidhjes së antigjenit të IgE, qelizat marrin një sinjal për të sekretuar substanca biologjikisht aktive (histamine, serotonin), të cilat janë kryesisht përgjegjëse për zhvillimin e reaksionit inflamator dhe për shfaqjen e reaksioneve alergjike si p.sh. astma, urtikarie, ethet e barit.
Imunoglobulinat D
Imunoglobulinat D gjenden në sasi shumë të vogla në serum; Zinxhirët e rëndë δ kanë një variabël dhe tre fusha konstante. IgD-të veprojnë si receptorë për limfocitet B, funksionet e tjera janë ende të panjohura. Ndërveprimi i antigjeneve specifike me receptorët në sipërfaqen e limfociteve B (IgD) çon në transmetimin e këtyre sinjaleve në qelizë dhe aktivizimin e mekanizmave që sigurojnë përhapjen e një kloni të caktuar limfocitar.
TEMA 1.7. VETITË FIZIKE DHE KIMIKE TË PROTEINAVE DHE METODAT PËR NDARJEN E TYRE
1. Proteinat individuale ndryshojnë në vetitë fizike dhe kimike:
Forma e molekulave;
pesha molekulare;
Ngarkesa totale, vlera e së cilës varet nga raporti i grupeve anionike dhe kationike të aminoacideve;
Raporti i radikaleve aminoacide polare dhe jopolare në sipërfaqen e molekulave;
Shkallët e rezistencës ndaj agjentëve të ndryshëm denatyrues.
2. Tretshmëria e proteinave varet mbi vetitë e proteinave të listuara më sipër, si dhe në përbërjen e mjedisit në të cilin proteina është tretur (vlerat e pH, përbërja e kripës, temperatura, prania e substancave të tjera organike që mund të ndërveprojnë me proteinën). Sasia e ngarkesës së molekulave të proteinave është një nga faktorët që ndikon në tretshmërinë e tyre. Kur ngarkesa në pikën izoelektrike humbet, proteinat grumbullohen dhe precipitojnë më lehtë. Kjo është veçanërisht tipike për proteinat e denatyruara, në të cilat radikalet hidrofobike të aminoacideve shfaqen në sipërfaqe.
Në sipërfaqen e një molekule proteine ka radikale aminoacide të ngarkuara pozitivisht dhe negativisht. Numri i këtyre grupeve dhe për rrjedhojë ngarkesa totale e proteinave varet nga pH e mjedisit, d.m.th. raporti i përqendrimeve të grupeve H+ - dhe OH -. Në një mjedis acid Një rritje në përqendrimin e H+ çon në shtypjen e disociimit të grupeve karboksil -COO - + H+ > - COOH dhe një ulje të ngarkesës negative të proteinave. Në një mjedis alkalik, lidhja e tepërt OH - nga protonet e formuara gjatë shpërbërjes së grupeve amino -NH 3 + + OH - - NH 2 + H 2 O me formimin e ujit, çon në një ulje të ngarkesës pozitive të proteinave. . Vlera e pH në të cilën një proteinë ka një ngarkesë neto zero quhet pika izoelektrike (IEP). Në IET, numri i grupeve të ngarkuara pozitivisht dhe negativisht është i njëjtë, d.m.th. proteina është në gjendje izoelektrike.
3. Ndarja e proteinave individuale. Karakteristikat e strukturës dhe funksionimit të trupit varen nga grupi i proteinave të sintetizuara në të. Studimi i strukturës dhe vetive të proteinave është i pamundur pa i izoluar ato nga qeliza dhe pa i pastruar nga proteinat e tjera dhe molekulat organike. Fazat e izolimit dhe pastrimit të proteinave individuale:
Shkatërrimi i qelizave indi që studiohet dhe përftohet një homogjen.
Ndarja e homogjenatit në fraksione me centrifugim, duke marrë një fraksion bërthamor, mitokondrial, citosolik ose fraksion tjetër që përmban proteinën e dëshiruar.
Denatyrim termik selektiv- Ngrohja afatshkurtër e një solucioni proteinik, gjatë së cilës mund të hiqen disa nga papastërtitë e proteinave të denatyruara (nëse proteina është relativisht e qëndrueshme ndaj nxehtësisë).
Kriposja. Proteina të ndryshme precipitojnë në përqendrime të ndryshme kripe në tretësirë. Duke rritur gradualisht përqendrimin e kripës, është e mundur të përftohen një numër fraksionesh të veçanta me një përmbajtje mbizotëruese të proteinave të izoluara në njërën prej tyre.
Sulfati i amonit përdoret më shpesh për fraksionimin e proteinave. Proteinat me tretshmëri më të vogël precipitojnë në përqendrime të ulëta kripërash. Filtrimi me xhel
- një metodë e shoshitjes së molekulave nëpër granula të fryra Sephadex (zinxhirë polisakarid tredimensionale të dekstranit që kanë pore). Shpejtësia me të cilën proteinat kalojnë nëpër një kolonë të mbushur me Sephadex do të varet nga pesha e tyre molekulare: sa më e vogël të jetë masa e molekulave të proteinave, aq më lehtë ato depërtojnë në granula dhe qëndrojnë atje sa më e madhe të jetë masa, aq më shpejt ato elustrojnë kolonë. Ultracentrifugimi
- një metodë që përfshin vendosjen e proteinave në një tub centrifuge në rotorin e një ultracentrifuge. Kur rotori rrotullohet, shkalla e sedimentimit të proteinave është proporcionale me peshën e tyre molekulare: fraksionet e proteinave më të rënda janë të vendosura më afër fundit të tubit të provës, ato më të lehta - më afër sipërfaqes. Elektroforeza
- një metodë e bazuar në ndryshimet në shpejtësinë e lëvizjes së proteinave në një fushë elektrike. Kjo vlerë është proporcionale me ngarkesën e proteinave. Elektroforeza e proteinave kryhet në letër (në këtë rast, shpejtësia e lëvizjes së proteinave është proporcionale vetëm me ngarkesën e tyre) ose në një xhel poliakrilamid me një madhësi të caktuar pore (shpejtësia e lëvizjes së proteinave është proporcionale me ngarkesën dhe peshën molekulare të tyre) .- një metodë fraksionimi e bazuar në lidhjen e grupeve të jonizuara të proteinave me grupe të ngarkuara në mënyrë të kundërt të rrëshirave të shkëmbimit të joneve (materiale polimerike të patretshme). Forca e lidhjes së proteinës me rrëshirën është proporcionale me ngarkesën e proteinës. Proteinat e absorbuara në polimerin e shkëmbimit të joneve mund të lahen me përqendrim në rritje të tretësirave të NaCl; Sa më i ulët të jetë ngarkesa e proteinave, aq më i ulët është përqendrimi i NaCl që kërkohet për të larë proteinën e lidhur me grupet jonike të rrëshirës.
Kromatografia e afinitetit- metoda më specifike për izolimin e proteinave individuale Një ligand i një proteine është i lidhur në mënyrë kovalente me një polimer inert. Kur një tretësirë proteine kalon nëpër një kolonë me një polimer, për shkak të lidhjes plotësuese të proteinës me ligand, vetëm proteina specifike për një ligand të caktuar absorbohet në kolonë.
Dializa- një metodë e përdorur për të hequr komponimet me peshë të ulët molekulare nga një zgjidhje e proteinave të izoluara. Metoda bazohet në paaftësinë e proteinave për të kaluar nëpër një membranë gjysmë të përshkueshme, ndryshe nga substancat me peshë të ulët molekulare. Përdoret për të pastruar proteinat nga papastërtitë me molekulare të ulët, për shembull, kripërat pas kriposjes.
DETYRA PËR PUNË JASHTËMËSIMORE
1. Plotësoni tabelën. 1.4.
Tabela 1.4. Analizë krahasuese e strukturës dhe funksioneve të proteinave të lidhura - mioglobinës dhe hemoglobinës
a) mbani mend strukturën e qendrës aktive të Mb dhe Hb. Çfarë roli luajnë radikalet hidrofobike të aminoacideve në formimin e qendrave aktive të këtyre proteinave? Përshkruani strukturën e qendrës aktive të Mb dhe Hb dhe mekanizmat e lidhjes së ligandëve me të. Çfarë roli luajnë mbetjet e tij F 8 dhe His E 7 në funksionimin e qendrës aktive të Mb dhe Hb?
b) çfarë veti të reja në krahasim me mioglobinën monomerike ka proteina oligomerike e lidhur ngushtë, hemoglobina? Shpjegoni rolin e ndryshimeve bashkëpunuese në konformimin e protomerëve në molekulën e hemoglobinës, efektin e përqendrimeve të CO 2 dhe protonit në afinitetin e hemoglobinës për oksigjenin, si dhe rolin e 2,3-BPG në rregullimin alosterik të funksionit të Hb. .
2. Karakterizoni kaperonët molekularë, duke i kushtuar vëmendje marrëdhënies midis strukturës dhe funksionit të tyre.
3. Cilat proteina ndahen në familje? Duke përdorur shembullin e familjes së imunoglobulinave, identifikoni karakteristika të ngjashme strukturore dhe funksione të ngjashme të proteinave të kësaj familjeje.
4. Proteinat individuale të pastruara shpesh kërkohen për qëllime biokimike dhe mjekësore. Shpjegoni se në cilat veti fiziko-kimike të proteinave bazohen metodat e përdorura për ndarjen dhe pastrimin e tyre.
DETYRAT E VETËKONTROLLIT
1. Zgjidhni përgjigjet e sakta.
Funksionet e hemoglobinës:
A. Transporti i O 2 nga mushkëritë në inde B. Transporti i H + nga indet në mushkëri
B. Mbajtja e një pH konstant të gjakut D. Transporti i CO 2 nga mushkëritë në inde
D. Transporti i CO 2 nga indet në mushkëri
2. Zgjidhni përgjigjet e sakta. Ligandα -protomeri Hb është: A. Heme
B. Oksigjen
B. CO G. 2,3-BPG
D. β-protomer
3. Zgjidhni përgjigjet e sakta.
Hemoglobina në krahasim me mioglobinën:
A. Ka një strukturë kuaternare
B. Struktura dytësore përfaqësohet vetëm nga α-helika
B. I përket proteinave komplekse
D. Ndërvepron me një ligand alosterik D. Lidhur në mënyrë kovalente me hemin
4. Zgjidhni përgjigjet e sakta.
Afiniteti i Hb për O2 zvogëlohet:
A. Kur shtohet një molekulë O 2 B. Kur hiqet një molekulë O 2
B. Kur ndërveproni me 2,3-BPG
D. Kur lidhet me protomerët H + D. Kur ulet përqendrimi i 2,3-BPG
5. Ndeshje.
Llojet HB karakterizohen nga:
A. Në formë deoksi formon agregate fibrilare B. Përmban dy zinxhirë α dhe dy δ
B. Forma mbizotëruese e Hb në eritrocitet e të rriturve D. Përmban hem me Fe+3 në qendrën aktive.
D. Përmban dy zinxhirë α- dhe dy γ 1. HbA 2.
6. Ndeshje.
Ligandët e Hb:
A. Lidhet me Hb në qendrën alosterike
B. Ka një afinitet shumë të lartë për vendin aktiv të Hb
B. Duke u bashkuar, rrit afinitetin e Hb për O 2 G. Oksidon Fe+ 2 në Fe + 3
D. Formon një lidhje kovalente me hisF8
7. Zgjidhni përgjigjet e sakta.
Chaperones:
A. Proteinat e pranishme në të gjitha pjesët e qelizës
B. Sinteza rritet nën stres
B. Merrni pjesë në hidrolizën e proteinave të denatyruara
D. Merrni pjesë në ruajtjen e konformacionit vendas të proteinave
D. Krijojnë organele në të cilat formohet konformacioni i proteinave.
8. Ndeshje. Imunoglobulinat:
A. Forma sekretore është pentamerike.
B. Klasa Ig që depërton në barrierën placentare
B. Ig - receptor i mastociteve
D. Klasa kryesore e Ig e pranishme në sekrecionet e qelizave epiteliale. D. Receptori i limfociteve B, aktivizimi i të cilit siguron proliferimin e qelizave
9. Zgjidhni përgjigjet e sakta.
Imunoglobulinat E:
A. Prodhohen nga makrofagët B. Kanë zinxhirë ε të rëndë.
B. Të ngulitura në membranën e limfociteve T
D. Veprojnë si receptorë të antigjenit të membranës në mastocitet dhe bazofilet
D. Përgjegjës për reaksionet alergjike
10. Zgjidhni përgjigjet e sakta.
Metoda për ndarjen e proteinave bazohet në ndryshimet në peshën e tyre molekulare:
A. Filtrimi me xhel
B. Ultracentrifugimi
B. Elektroforeza me xhel poliakrilamid D. Kromatografia e shkëmbimit të joneve
D. Kromatografia e afinitetit
11. Zgjidhni përgjigjen e saktë.
Metoda për ndarjen e proteinave bazohet në ndryshimet në tretshmërinë e tyre në ujë:
A. Filtrimi me xhel B. Kriposja
B. Kromatografia e shkëmbimit të joneve D. Kromatografia e afinitetit
D. Elektroforeza në xhel poliakrilamid
STANDARDET E PËRGJIGJEVE NË "DETYRAT E VETËKontrollimit"
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
TERMAT DHE KONCEPTET THEMELORE
1. Proteinat oligomerike, protomeri, struktura kuaternare e proteinave
2. Ndryshimet bashkëpunuese në konformacionin e protomerit
3. Efekti Bohr
4. Rregullimi alosterik i funksioneve proteinike, qendra alosterike dhe efektori alosterik
5. Kaperone molekulare, proteina të goditjes nga nxehtësia
6. Familjet e proteinave (proteazat serine, imunoglobulinat)
7. Marrëdhënia IgM-, G-, E-, A-strukturë-funksion
8. Ngarkesa totale e proteinave, pika izoelektrike e proteinave
9. Elektroforeza
10. Kriposja
11. Filtrim me xhel
12. Kromatografia e shkëmbimit të joneve
13. Ultracentrifugimi
14. Kromatografia e afinitetit
15. Elektroforeza e proteinave të plazmës së gjakut
DETYRA PËR PUNËN NË KLASË
1. Krahasoni varësinë e shkallëve të ngopjes së hemoglobinës (Hb) dhe mioglobinës (Mb) me oksigjenin nga presioni i tij i pjesshëm në inde
 Oriz. 1.34. Varësia e ngopjes Mv dheNHoksigjen nga presioni i pjesshëm i tij
Oriz. 1.34. Varësia e ngopjes Mv dheNHoksigjen nga presioni i pjesshëm i tij
Ju lutemi vini re se forma e kurbave të ngopjes së oksigjenit të proteinave është e ndryshme: për mioglobinën - një hiperbolë, për hemoglobinën - një formë sigmoide.
1. Krahasoni vlerat e presionit të pjesshëm të oksigjenit në të cilin Mb dhe Hb janë të ngopura me O 2 me 50%. Cila nga këto proteina ka një afinitet më të lartë për O 2?
2. Cilat veçori strukturore të Mb përcaktojnë afinitetin e tij të lartë për O 2?
3. Cilat veçori strukturore të HB e lejojnë atë të lëshojë O2 në kapilarët e indeve në pushim (në një presion pjesor relativisht të lartë të O2) dhe të rrisë ndjeshëm këtë lëshim në muskujt që punojnë? Cila veti e proteinave oligomerike e jep këtë efekt?
4. Llogaritni sa sasi O 2 (në%) i jep hemoglobinës së oksigjenuar për muskujt që pushojnë dhe punojnë?
5. të nxjerrë përfundime për marrëdhënien midis strukturës së një proteine dhe funksionit të saj.
2. Sasia e oksigjenit të çliruar nga hemoglobina në kapilarë varet nga intensiteti i proceseve katabolike në inde (efekti Bohr). Si e rregullojnë ndryshimet në metabolizmin e indeve afinitetin e Hb për O2? Efekti i CO 2 dhe H+ në afinitetin e Hb për O 2
 1. përshkruani efektin Bohr.
1. përshkruani efektin Bohr.
2. në çfarë drejtimi vazhdon procesi i paraqitur në diagram:
a) në kapilarët e mushkërive;
b) në kapilarët e indeve?
3. Cila është rëndësia fiziologjike e efektit Bohr?
4. Pse ndërveprimi i Hb me H+ në vende të largëta nga hemi e ndryshon afinitetin e proteinës për O 2?
3. Afiniteti i Hb për O2 varet nga përqendrimi i ligandit të tij - 2,3-bisfosfoglicerati, i cili është një rregullator alosterik i afinitetit të Hb për O2.
Pse ndërveprimi i ligandit në një vend të largët nga zona aktive ndikon në funksionin e proteinave? Si e rregullon 2,3-BPG afinitetin e Hb për O2? Për të zgjidhur problemin, përgjigjuni pyetjeve të mëposhtme:
1. ku dhe nga çfarë sintetizohet 2.3-bisfosfoglicerati (2,3-BPG)? Shkruani formulën e saj, tregoni ngarkesën e kësaj molekule.
2. Me cilën formë të hemoglobinës (oksi apo deoksi) ndërvepron BPG dhe pse? Në cilën pjesë të molekulës së Hb ndodh ndërveprimi?
3. në çfarë drejtimi ecën procesi i paraqitur në diagram?
a) në kapilarët e indeve;
b) në kapilarët e mushkërive?
4. ku përqendrimi i kompleksit duhet të jetë më i lartë
Nv-2,3-BFG:
a) në kapilarët e muskujve në pushim,
b) në kapilarët e muskujve që punojnë (siguroi të njëjtin përqendrim të BPG në eritrocite)?
4. 5. Si do të ndryshojë afiniteti i HB për oksigjenin kur një person përshtatet me kushtet e lartësisë së madhe, nëse rritet përqendrimi i BPG në eritrocite? Cila është rëndësia fiziologjike e këtij fenomeni?
5. Shkatërrimi i 2,3-BPG gjatë ruajtjes së gjakut të konservuar dëmton funksionet e HB. Si do të ndryshojë afiniteti i HB për O 2 në gjakun e konservuar nëse përqendrimi i 2,3-BPG në eritrocite mund të ulet nga 8 në 0,5 mmol/l. A është e mundur transfuzioni i një gjaku të tillë te pacientët e sëmurë rëndë nëse përqendrimi i 2,3-BPG rikthehet jo më herët se pas tre ditësh? A është e mundur të rivendosni funksionet e qelizave të kuqe të gjakut duke shtuar 2,3-BPG në gjak?
Mos harroni strukturën e molekulave më të thjeshta të imunoglobulinës. Çfarë roli luajnë imunoglobulinat në sistemin imunitar? Pse Ig-të shpesh quhen bivalente? Si lidhet struktura e Ig-ve me funksionin e tyre? (Përshkruani duke përdorur një shembull të një klase të imunoglobulinave.)
6. Vetitë fiziko-kimike të proteinave dhe metodat e ndarjes së tyre.
Si ndikon ngarkesa neto e një proteine në tretshmërinë e saj?
a) përcaktoni ngarkesën totale të peptidit në pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis<7, рН <<7?
b) si do të ndryshojë ngarkesa e këtij peptidi në pH >7, pH
c) cila është pika izoelektrike e një proteine (IEP) dhe në çfarë mjedisi ndodhet ajo?
IET i këtij peptidi?
7. d) në çfarë vlere pH do të vërehet tretshmëria më e ulët e këtij peptidi.
8. Filtrimi me xhel përdoret për të ndarë proteinat individuale. Një përzierje që përmban proteina A, B, C me pesha molekulare të barabartë me 160,000, 80,000 dhe 60,000, respektivisht, u analizua me anë të filtrimit me xhel (Fig. 1.35). Granulat e xhelit të fryrë janë të përshkueshme ndaj proteinave me një peshë molekulare më të vogël se 70,000? Cili grafik pasqyron saktë rezultatet e fraksionimit? Tregoni rendin në të cilin proteinat A, B dhe C lirohen nga kolona.
 Oriz. 1.35. Përdorimi i filtrimit me xhel për ndarjen e proteinave
Oriz. 1.35. Përdorimi i filtrimit me xhel për ndarjen e proteinave
9. Në Fig. 1.36, A tregon një diagram të elektroforezës në letër të proteinave të serumit të gjakut të një personi të shëndetshëm. Sasitë relative të fraksioneve proteinike të përftuara me këtë metodë janë: albuminat 54-58%, α1-globulina 6-7%, α2-globulina 8-9%, β-globulina 13%, γ-globulina 11-12%.
 Oriz. 1.36 Elektroforeza në letër e proteinave të plazmës së gjakut të një personi të shëndetshëm (A) dhe një pacienti (B)
Oriz. 1.36 Elektroforeza në letër e proteinave të plazmës së gjakut të një personi të shëndetshëm (A) dhe një pacienti (B)
I - γ-globulina; II - β-globulina; III -α 2 -globulina; IV -α 2 -globulina; V - albuminat
Shumë sëmundje shoqërohen me ndryshime sasiore në përbërjen e proteinave të serumit (disproteinemia). Natyra e këtyre ndryshimeve merret parasysh kur bëhet një diagnozë dhe vlerësohet ashpërsia dhe faza e sëmundjes.
Duke përdorur të dhënat e dhëna në tabelë. 1.5, bëni një supozim për sëmundjen, e cila karakterizohet nga profili elektroforetik i paraqitur në Fig. 1.36.
Tabela 1.5. Ndryshimet në përqendrimin e proteinave të serumit në patologji
Një proteinë është një sekuencë e aminoacideve të lidhura me njëri-tjetrin me lidhje peptide.
Është e lehtë të imagjinohet se numri i aminoacideve mund të jetë i ndryshëm: nga një minimum prej dy në çdo vlerë të arsyeshme. Biokimistët kanë rënë dakord që nëse numri i aminoacideve nuk kalon 10, atëherë një përbërje e tillë quhet peptid; nëse ka 10 ose më shumë aminoacide - një polipeptid. Polipeptidet që janë në gjendje të formojnë spontanisht dhe të mbajnë një strukturë të caktuar hapësinore, të quajtur konformacion, klasifikohen si proteina. Stabilizimi i një strukture të tillë është i mundur vetëm kur polipeptidet arrijnë një gjatësi të caktuar (më shumë se 40 aminoacide prandaj, polipeptidet me një peshë molekulare prej më shumë se 5000 Da zakonisht konsiderohen proteina). (1Da është e barabartë me 1/12 e një izotopi të karbonit). Vetëm duke pasur një strukturë të caktuar hapësinore (strukturë amtare) një proteinë mund të kryejë funksionet e saj.
Madhësia e proteinës mund të matet në dalton (pesha molekulare), shpesh për shkak të madhësisë relativisht të madhe të molekulës në njësitë e saj të prejardhura, kilodaltons (kDa). Proteinat e majave, mesatarisht, përbëhen nga 466 aminoacide dhe kanë një peshë molekulare prej 53 kDa. Proteina më e madhe e njohur aktualisht, titina, është një komponent i sarkomerëve të muskujve; Pesha molekulare e izoformave të ndryshme të saj varion nga 3000 në 3700 kDa dhe përbëhet nga 38138 aminoacide (në muskulin solius të njeriut).
Struktura e proteinave
Struktura tredimensionale e një proteine formohet gjatë procesit të palosjes. e palosshme -"paloset") Një strukturë tre-dimensionale formohet si rezultat i ndërveprimit të strukturave në nivele më të ulëta.
Ekzistojnë katër nivele të strukturës së proteinave:
Struktura primare- sekuenca e aminoacideve në një zinxhir polipeptid.
Struktura dytësore- kjo është vendosja në hapësirë e seksioneve individuale të zinxhirit polipeptid.
Më poshtë janë llojet më të zakonshme të strukturës dytësore të proteinave:
α-helika- kthesa të dendura rreth boshtit të gjatë të molekulës, një kthesë përbëhet nga 3.6 mbetje aminoacide, dhe hapi i spirales është 0.54 nm (0.15 nm për mbetje aminoacide), spiralen stabilizohet nga lidhjet hidrogjenore midis H dhe O e grupeve peptide të vendosura larg njëri-tjetrit nga 4 mbetje aminoacide. Spiralja është ndërtuar ekskluzivisht nga një lloj stereoizomeri i aminoacideve (L). Edhe pse mund të jetë i majtë ose i djathtë, djathtasja është mbizotëruese në proteina. Helix është ndërprerë nga ndërveprimet elektrostatike të acidit glutamik, lizinës dhe argininës. Mbetjet e asparaginës, serinës, treoninës dhe leucinës të vendosura afër njëra-tjetrës mund të ndërhyjnë në mënyrë sterike në formimin e spirales, Mbetjet e prolinës shkaktojnë lakimin e zinxhirit dhe gjithashtu prishin strukturën e α-spiralës.
shtresa të palosura β- disa zinxhirë polipeptidësh zigzag, në të cilët formohen lidhje hidrogjenore midis aminoacideve ose zinxhirëve të ndryshëm proteinash relativisht të largët nga njëri-tjetri (0,347 nm për mbetje aminoacide) në strukturën parësore, dhe jo të vendosura ngushtë, siç është rasti në një spirale α. . Këto zinxhirë zakonisht kanë skajet e tyre N-terminale në drejtime të kundërta (orientim antiparalel). Përmasat e vogla të grupeve anësore të aminoacideve janë të rëndësishme për formimin e fletëve të β-fletave zakonisht mbizotërojnë glicina dhe alanina;

Palosja e proteinave në një fletë të palosur β
Strukturat e çrregulluara janë një rregullim i çrregullt i një zinxhiri proteinik në hapësirë.
Struktura hapësinore e secilës proteinë është individuale dhe përcaktohet nga struktura e saj primare. Megjithatë, një krahasim i konformacioneve të proteinave me struktura dhe funksione të ndryshme zbuloi praninë e kombinimeve të ngjashme të elementeve të strukturës dytësore në to. Ky rend specifik i formimit të strukturave dytësore quhet strukturë supersekondare e proteinave. Struktura supersekondare formohet për shkak të ndërveprimeve ndërradikale.
Disa kombinime karakteristike të α-helikave dhe strukturave β shpesh quhen "motive strukturore". Ata kanë emra specifikë: “α-helix-turn-α-helix”, “a/β-fuci strukturë”, “zinxher leucine”, “gisht zink” etj.
Struktura terciare- Kjo është një mënyrë për të vendosur të gjithë zinxhirin polipeptid në hapësirë. Së bashku me α-helika, fletët me palosje β dhe strukturat supersekondare, struktura terciare zbulon një konformacion të çrregullt që mund të zërë një pjesë të konsiderueshme të molekulës.

Paraqitja skematike e palosjes së proteinave në strukturë terciare.
Struktura kuaternare ndodh në proteinat që përbëhen nga disa vargje polipeptide (nënnjësi, protomere ose monomere), kur strukturat terciare të këtyre nënnjësive kombinohen. Për shembull, molekula e hemoglobinës përbëhet nga 4 nënnjësi. Formacionet supramolekulare kanë një strukturë kuaternare - komplekse multienzimash, të cilat përbëhen nga disa molekula enzimash dhe koenzimash (piruvat dehidrogjenaza), dhe izoenzima (laktat dehidrogjenaza - LDH, kreatine fosfokinaza - CPK).
Pra. Struktura hapësinore nuk varet nga gjatësia e zinxhirit polipeptid, por nga sekuenca e mbetjeve të aminoacideve specifike për secilën proteinë, si dhe nga radikalet anësore karakteristike të aminoacideve përkatëse. Struktura hapësinore tredimensionale ose konformimi i makromolekulave të proteinave formohet kryesisht nga lidhjet hidrogjenore, ndërveprimet hidrofobike midis radikaleve anësore jopolare të aminoacideve dhe ndërveprimet jonike midis grupeve anësore të ngarkuara në mënyrë të kundërt të mbetjeve të aminoacideve. Lidhjet e hidrogjenit luajnë një rol të madh në formimin dhe mirëmbajtjen e strukturës hapësinore të makromolekulës së proteinës.
Sa i përket ndërveprimeve hidrofobike, ato lindin si rezultat i kontaktit midis radikalëve jopolare që nuk janë në gjendje të thyejnë lidhjet hidrogjenore midis molekulave të ujit, të cilat zhvendosen në sipërfaqen e globulës së proteinës. Ndërsa sinteza e proteinave vazhdon, grupet kimike jopolare grumbullohen brenda globulës dhe ato polare detyrohen të dalin në sipërfaqen e saj. Kështu, një molekulë proteine mund të jetë neutrale, e ngarkuar pozitivisht ose e ngarkuar negativisht, në varësi të pH-së së tretësit dhe grupeve jonike në proteinë. Për më tepër, konformimi i proteinave ruhet nga lidhjet kovalente S-S të formuara midis dy mbetjeve të cisteinës. Si rezultat i formimit të strukturës amtare të proteinës, shumë atome të vendosura në pjesë të largëta të zinxhirit polipeptid afrohen dhe, duke ndikuar njëri-tjetrin, fitojnë veti të reja që mungojnë në aminoacide individuale ose polipeptide të vogla.
Është e rëndësishme të kuptohet se palosja - palosja e proteinave (dhe biomakromolekulave të tjera) nga një konformacion i shpalosur në një formë "vendase" - është një proces fizik dhe kimik, si rezultat i të cilit proteinat në "habitatin" e tyre natyror (zgjidhje, citoplazma ose membrana) fitojnë karakteristika karakteristike vetëm të tyre shtrirjes dhe funksioneve hapësinore.
Qelizat përmbajnë një numër proteinash joaktive katalitike, të cilat megjithatë japin një kontribut të madh në formimin e strukturave të proteinave hapësinore. Këta janë të ashtuquajturit chaperone. Kaperonët ndihmojnë në montimin e saktë të konformacionit të proteinave tredimensionale duke formuar komplekse të kthyeshme jokovalente me zinxhirin polipeptid pjesërisht të palosur, ndërsa njëkohësisht pengojnë lidhjet e keqformuara që çojnë në formimin e strukturave të proteinave funksionalisht joaktive. Lista e funksioneve karakteristike për kaperonet përfshin mbrojtjen e globulave të shkrirë (pjesërisht të palosura) nga grumbullimi, si dhe transferimin e proteinave të saposintetizuara në lokacione të ndryshme qelizore.
Kaperonet janë kryesisht proteina të goditjes së nxehtësisë, sinteza e të cilave rritet ndjeshëm nën ndikimet stresuese të temperaturës, kjo është arsyeja pse ato quhen edhe hsp (proteinat e goditjes nga nxehtësia). Familjet e këtyre proteinave gjenden në qelizat mikrobike, bimore dhe shtazore. Klasifikimi i kaperonëve bazohet në peshën e tyre molekulare, e cila varion nga 10 në 90 kDa. Ato janë proteina që ndihmojnë në formimin e strukturës tre-dimensionale të proteinave. Kaperonët e mbajnë zinxhirin polipeptid të saposintetizuar në një gjendje të shpalosur, duke e penguar atë të paloset në një formë të ndryshme nga ajo vendase, dhe sigurojnë kushte për strukturën e vetme të saktë të proteinës vendase.
Gjatë palosjes së proteinave, disa konformacione të molekulës refuzohen në fazën e globulës së shkrirë. Degradimi i molekulave të tilla nis nga proteina ubiquitin.
Degradimi i proteinave përmes rrugës së ubiquitin përfshin dy faza kryesore:
1) lidhja kovalente e ubiquitinit me proteinën që do të degradohet përmes një mbetjeje lizinë, prania e një etikete të tillë në proteinë është sinjali kryesor i klasifikimit që drejton konjugatët që rezultojnë te proteazomet, në shumicën e rasteve, disa molekula ubiquitine, të cilat janë të organizuara në formën e rruazave në një varg, janë ngjitur në proteinë;
2) hidroliza e proteinave nga proteazoma (funksioni kryesor i proteazomës është degradimi proteolitik i proteinave të panevojshme dhe të dëmtuara në peptide të shkurtra). Ubiquitin quhet me të drejtë "shenja e vdekjes" për proteinat.
Dom?n ketri? - një element i strukturës terciare të një proteine, e cila është një nënstrukturë mjaft e qëndrueshme dhe e pavarur e proteinës, palosja e së cilës ndodh në mënyrë të pavarur nga pjesët e tjera. Një domen zakonisht përfshin disa elementë të strukturës dytësore. Domene strukturore të ngjashme gjenden jo vetëm në proteinat e lidhura (për shembull, në hemoglobinat e kafshëve të ndryshme), por edhe në proteina krejtësisht të ndryshme. Një proteinë mund të ketë domene të shumta, dhe këto rajone mund të kryejnë funksione të ndryshme në të njëjtën proteinë. Disa enzima dhe të gjitha imunoglobulinat kanë një strukturë domeni. Proteinat me zinxhirë të gjatë polipeptidikë (më shumë se 200 mbetje aminoacide) shpesh krijojnë struktura domenesh.