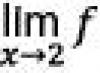La protéine catalase remplit une fonction dans la cellule ;
Classe
1. Les éléments les plus courants dans les cellules des organismes vivants sont :
2. L'azote en tant qu'élément est inclus dans :
3. L'hydrogène en tant qu'élément est inclus dans :
4. À quel niveau d'organisation n'y a-t-il aucune différence entre le monde organique et inorganique ?
5.Plus d’eau est contenue dans les cellules :
6. L’eau est la base de la vie :
7. Les substances hautement solubles dans l'eau sont appelées :
8. Les composés hydrophobes de la cellule comprennent :
9. Les glucides monosaccharides comprennent :
10. Les glucides et les polysaccharides comprennent :
11. Les principales fonctions des graisses dans la cellule :
12. Les protéines sont des biopolymères avec des monomères, qui sont :
13. Les acides aminés diffèrent :
14. La composition des molécules protéiques comprend :
15. La structure d'une molécule protéique, qui est déterminée par la séquence de résidus d'acides aminés :
16. La structure secondaire d'une protéine est associée à :
17. Il existe un certain lien entre le premier et le deuxième concepts de la tâche Trouvez ce mot Cellule : chloroplaste = plante : _______________.
18. Les protéines structurelles les moins fortes sont :
20. Lorsqu'une protéine est incomplètement dénaturée, la structure est d'abord détruite :
21. Les monomères des molécules d'ADN sont :
22. Les nucléotides d'ADN sont constitués de :
23. La composition des nucléotides d'ADN diffère les unes des autres par leur contenu :
24. Les nucléotides d'ADN contiennent des bases azotées :
25. Les nucléotides d'ARN sont constitués de :
26. Molécules dont l'oxydation libère beaucoup d'énergie :
27. Les éléments les plus courants dans les cellules des organismes vivants sont :
28. Le carbone en tant qu'élément est inclus dans :
29. Fonctions de l'eau dans une cellule :
30. Les glucides monosaccharides comprennent :
31. Les glucides et les polysaccharides comprennent :
32. La molécule d'ADN contient des résidus :
33. Les produits de la réaction entre le glycérol et les acides gras supérieurs sont :
34. Les graisses et les huiles ont les propriétés suivantes par rapport à l'eau :
35. Les protéines sont :
36. Dans les solutions aqueuses, les acides aminés présentent les propriétés suivantes :
37. La structure primaire d'une protéine est déterminée par :
38. La structure primaire d'une protéine est soutenue par des liaisons :
39. Les enzymes remplissent les fonctions suivantes :
40. L'activité biologique d'une protéine est déterminée par sa structure :
41. Molécules qui se décomposent le plus facilement dans une cellule pour libérer de l'énergie :
42. Les monomères des molécules d'acide nucléique sont :
43. Les nucléotides d'une molécule d'ADN contiennent des bases azotées :
44. Contient plus de glucides :
45. Les nucléotides d'une molécule d'ARN contiennent des bases azotées :
46. Les graisses sont solubles :
47. Une molécule d'une substance constituée de nucléotides et ayant la forme d'un fil simple brin :
48. Les plus grosses molécules parmi les acides nucléiques sont :
49. Les sels K sont importants pour le corps car :
50. La science qui étudie le fonctionnement des organismes s'appelle :
51. La capacité d'un mode de nutrition chimioautotrophe est caractéristique de :
52. Substances qui servent d'accumulateurs d'énergie biologique universels dans la cellule :
53. Dans une molécule d'ADN, le nombre de nucléotides contenant de la cytosine représente 15 % du nombre total. Quel est le pourcentage de nucléotides contenant de l'adénine dans cette molécule ?
54. Le résidu d'acide aminé d'une protéine est codé :
55. La séquence de nucléotides dans l'une des chaînes d'ADN complémentaires est AGA. Quelle est la séquence nucléotidique correspondante dans l’autre chaîne ? :
56. Les cellules fongiques, comme les cellules animales, manquent de :
57. Organites cellulaires responsables de son mouvement :
58. Les personnes suivantes ont leur propre ADN :
59. Parmi les réponses proposées, choisissez l'une des dispositions de la théorie cellulaire:
61. L'ATP est considérée comme la principale source d'énergie des cellules, car :
62. Le métabolisme se produit dans chaque cellule vivante et est :
63. Quelle est la principale source d'énergie qui assure la circulation des substances dans les écosystèmes ?
Les protéines (protéines) représentent 50 % de la masse sèche des organismes vivants.
Les protéines sont constituées d'acides aminés. Chaque acide aminé possède un groupe aminé et un groupe acide (carboxyle), dont l'interaction produit liaison peptidique C’est pourquoi les protéines sont également appelées polypeptides.
Structures protéiques
Primaire- une chaîne d'acides aminés liés par une liaison peptidique (forte, covalente). En alternant 20 acides aminés dans des ordres différents, vous pouvez créer des millions de protéines différentes. Si vous modifiez au moins un acide aminé dans la chaîne, la structure et les fonctions de la protéine changeront, c'est pourquoi la structure primaire est considérée comme la plus importante de la protéine.
Secondaire- spirale. Détenu par des liaisons hydrogène (faibles).
Tertiaire- globule (boule). Quatre types de liaisons : le disulfure (pont soufre) est fort, les trois autres (ionique, hydrophobe, hydrogène) sont faibles. Chaque protéine a sa propre forme de globule et ses fonctions en dépendent. Lors de la dénaturation, la forme du globule change, ce qui affecte le fonctionnement de la protéine.
Quaternaire- Toutes les protéines n'en ont pas. Elle est constituée de plusieurs globules reliés entre eux par les mêmes liaisons que dans la structure tertiaire. (Par exemple, l'hémoglobine.)
Dénaturation
Il s'agit d'un changement de forme d'un globule protéique provoqué par des influences extérieures (température, acidité, salinité, ajout d'autres substances, etc.)
- Si les effets sur la protéine sont faibles (changement de température de 1°), alors réversible dénaturation.
- Si l'impact est fort (100°), alors dénaturation irréversible. Dans ce cas, toutes les structures sauf la principale sont détruites.
Fonctions des protéines
Il y en a beaucoup, par exemple :
- Enzymatique (catalytique)- les protéines enzymatiques accélèrent les réactions chimiques du fait que le centre actif de l'enzyme épouse la forme de la substance, comme la clé d'une serrure (spécificité).
- Construction (structurelle)- la cellule, outre l'eau, est constituée principalement de protéines.
- Protecteur- les anticorps combattent les pathogènes (immunité).
Choisissez-en une, l'option la plus correcte. La structure secondaire d'une molécule protéique a la forme
1) spirales
2) double hélice
3) balle
4) fils
Répondre
Choisissez-en une, l'option la plus correcte. Les liaisons hydrogène entre les groupes CO et NH dans la molécule protéique lui confèrent la forme hélicoïdale caractéristique de la structure
1) primaire
2) secondaire
3) tertiaire
4) quaternaire
Répondre
Choisissez-en une, l'option la plus correcte. Le processus de dénaturation d'une molécule protéique est réversible si les liaisons ne sont pas rompues
1) hydrogène
2) peptides
3) hydrophobe
4) disulfure
Répondre
Choisissez-en une, l'option la plus correcte. La structure quaternaire d'une molécule protéique se forme à la suite de l'interaction
1) sections d'une molécule protéique selon le type de liaisons S-S
2) plusieurs brins polypeptidiques formant une boule
3) sections d'une molécule de protéine dues aux liaisons hydrogène
4) globule protéique avec membrane cellulaire
Répondre
Établir une correspondance entre la caractéristique et la fonction de la protéine qu'elle remplit : 1) régulatrice, 2) structurelle
A) fait partie des centrioles
B) forme des ribosomes
B) est une hormone
D) forme les membranes cellulaires
D) modifie l'activité des gènes
Répondre
Choisissez-en une, l'option la plus correcte. La séquence et le nombre d'acides aminés dans une chaîne polypeptidique sont
1) structure primaire de l'ADN
2) structure protéique primaire
3) structure secondaire de l'ADN
4) structure secondaire de la protéine
Répondre
Choisissez trois options. Protéines chez l'homme et l'animal
1) servir de matériau de construction principal
2) sont décomposés dans les intestins en glycérol et en acides gras
3) sont formés d'acides aminés
4) dans le foie, ils sont convertis en glycogène
5) mettre en réserve
6) en tant qu'enzymes, ils accélèrent les réactions chimiques
Répondre
Choisissez-en une, l'option la plus correcte. La structure secondaire de la protéine, qui a la forme d'une hélice, est maintenue par des liaisons
1) peptide
2) ionique
3) hydrogène
4) covalent
Répondre
Choisissez-en une, l'option la plus correcte. Quelles liaisons déterminent la structure primaire des molécules protéiques
1) hydrophobe entre les radicaux d'acides aminés
2) hydrogène entre les brins polypeptidiques
3) peptide entre les acides aminés
4) hydrogène entre les groupes -NH- et -CO-
Répondre
Choisissez-en une, l'option la plus correcte. La structure primaire d'une protéine est formée par une liaison
1) hydrogène
2) macroergique
3) peptides
4) ionique
Répondre
Choisissez-en une, l'option la plus correcte. La formation de liaisons peptidiques entre les acides aminés dans une molécule protéique est basée sur
1) principe de complémentarité
2) insolubilité des acides aminés dans l'eau
3) solubilité des acides aminés dans l'eau
4) la présence de groupes carboxyle et amine en eux
Répondre
Les caractéristiques énumérées ci-dessous, sauf deux, sont utilisées pour décrire la structure et les fonctions de la matière organique représentée. Identifiez deux caractéristiques qui « disparaissent » de la liste générale et notez les numéros sous lesquels elles sont indiquées.
1) possède des niveaux structurels d’organisation de la molécule
2) fait partie des parois cellulaires
3) est un biopolymère
4) sert de matrice pour la traduction
5) se compose d'acides aminés
Répondre
Toutes les caractéristiques suivantes, sauf deux, peuvent être utilisées pour décrire les enzymes. Identifiez deux caractéristiques qui « disparaissent » de la liste générale et notez les numéros sous lesquels elles sont indiquées.
1) font partie des membranes cellulaires et des organites cellulaires
2) jouer le rôle de catalyseurs biologiques
3) avoir un centre actif
4) influencer le métabolisme, régulant divers processus
5) protéines spécifiques
Répondre

Regardez l'image d'un polypeptide et indiquez (A) son niveau d'organisation, (B) la forme de la molécule et (C) le type d'interaction qui maintient la structure. Pour chaque lettre, sélectionnez le terme ou le concept correspondant dans la liste fournie.
1) structure primaire
2) structure secondaire
3) structure tertiaire
4) interactions entre nucléotides
5) connexion métallique
6) interactions hydrophobes
7) fibrillaire
8) globulaire
Répondre

Regardez l'image d'un polypeptide. Indiquez (A) son niveau d'organisation, (B) les monomères qui le composent et (C) le type de liaisons chimiques entre eux. Pour chaque lettre, sélectionnez le terme ou le concept correspondant dans la liste fournie.
1) structure primaire
2) liaisons hydrogène
3) double hélice
4) structure secondaire
5) acide aminé
6) hélice alpha
7) nucléotide
8) liaisons peptidiques
Répondre
On sait que les protéines sont des polymères irréguliers de poids moléculaire élevé et strictement spécifiques à chaque type d’organisme. Sélectionnez trois énoncés dans le texte ci-dessous qui sont liés de manière significative à la description de ces caractéristiques et notez les numéros sous lesquels ils sont indiqués.
Répondre
(1) Les protéines contiennent 20 acides aminés différents liés par des liaisons peptidiques. (2) Les protéines ont différents nombres d'acides aminés et l'ordre de leur alternance dans la molécule. (3) Les substances organiques de faible poids moléculaire ont un poids moléculaire de 100 à 1 000. (4) Ce sont des composés intermédiaires ou des unités structurelles - des monomères. (5) De nombreuses protéines sont caractérisées par un poids moléculaire de plusieurs milliers à un million ou plus, en fonction du nombre de chaînes polypeptidiques individuelles dans la structure moléculaire unique de la protéine. (6) Chaque type d’organisme vivant possède un ensemble spécial et unique de protéines qui le distingue des autres organismes.
© D.V. Pozdniakov, 2009-2019
Les protéines sont des biopolymères dont les monomères sont des acides aminés. Acides aminés
sont des composés organiques de faible poids moléculaire contenant des groupes carboxyle (-COOH) et amine (-NH 2) qui sont liés au même atome de carbone. Une chaîne latérale est attachée à l'atome de carbone - un radical qui confère à chaque acide aminé certaines propriétés. La plupart des acides aminés ont un groupe carboxyle et un groupe amino ; ces acides aminés sont appelés neutre . Il existe cependant aussi acides aminés basiques - avec plus d'un groupe amino, ainsi que acide acides aminés
- avec plus d'un groupe carboxyle. Il existe environ 200 acides aminés connus dans les organismes vivants, mais seulement 20 d’entre eux se trouvent dans les protéines. Ce sont les soi-disant basique ou protéinogène
acides aminés.
- Selon le radical, les acides aminés basiques sont divisés en 3 groupes :
- Non polaire (alanine, méthionine, valine, proline, leucine, isoleucine, tryptophane, phénylalanine) ;
- Polaire non chargé (asparagine, glutamine, sérine, glycine, tyrosine, thréonine, cystéine) ;
Chargé (arginine, histidine, lysine - positivement ; acide aspartique et glutamique - négatif).
Les chaînes latérales d'acides aminés (radicaux) peuvent être hydrophobes ou hydrophiles et conférer les propriétés correspondantes aux protéines. irremplaçable. Ceux-ci incluent la lysine, la valine, la leucine, l'isoleucine, la thréonine, la phénylalanine, le tryptophane, la méthionine ; l'arginine et l'histidine sont essentielles pour les enfants.
En solution, les acides aminés peuvent agir à la fois comme acides et comme bases, c'est-à-dire qu'ils sont des composés amphotères. Le groupe carboxyle (-COOH) peut donner un proton, fonctionnant comme un acide, et le groupe amino (-NH 2) peut accepter un proton, présentant ainsi les propriétés d'une base.
Le groupe amino d’un acide aminé peut réagir avec le groupe carboxyle d’un autre acide aminé. La molécule résultante est dipeptide, et la liaison entre les atomes de carbone et d’azote est appelée liaison peptidique.
À une extrémité de la molécule dipeptide se trouve un groupe amino libre et à l’autre, un groupe carboxyle libre. Grâce à cela, le dipeptide peut s'attacher à lui-même d'autres acides aminés, formant ainsi des oligopeptides. Si de nombreux acides aminés (plus de 10) sont ainsi combinés, alors polypeptide.
Les peptides jouent un rôle important dans l'organisme. De nombreux aligopeptides sont des hormones. Il s'agit de l'ocytocine, de la vasopressine, de la thyrolibérine, de la thyrotropine, etc. Les oligopeptides comprennent également la bradykidine (peptide de la douleur) et certains opiacés (« médicaments naturels » de l'homme), qui remplissent la fonction de soulagement de la douleur. La prise de drogues détruit le système opiacé du corps, de sorte qu'un toxicomane sans dose de drogue éprouve une douleur intense - un « sevrage », qui est normalement soulagé par les opiacés.
Les oligopeptides comprennent certains antibiotiques (par exemple, la gramicidine S).
De nombreuses hormones (insuline, hormone adrénocorticotrope, etc.), antibiotiques (par exemple, gramicidine A), toxines (par exemple, toxine diphtérique) sont des polypeptides.
Les protéines sont des polypeptides d'un poids moléculaire supérieur à 10 000, dont la molécule contient de 50 à plusieurs milliers d'acides aminés.
Chaque protéine a sa propre structure spatiale particulière dans un certain environnement. Lors de la caractérisation de la structure spatiale (tridimensionnelle), on distingue quatre niveaux d'organisation des molécules protéiques.
Structure primaire- séquence d'acides aminés dans une chaîne polypeptidique. La structure primaire est spécifique à chaque protéine et est déterminée par l'information génétique, c'est-à-dire dépend de la séquence de nucléotides dans la section de la molécule d'ADN codant pour la protéine. Toutes les propriétés et fonctions des protéines dépendent de la structure primaire. Le remplacement d’un seul acide aminé dans des molécules protéiques ou la modification de leur disposition entraîne généralement une modification de la fonction des protéines. Étant donné que les protéines contiennent 20 types d'acides aminés, le nombre d'options pour leurs combinaisons dans la chaîne sexuelle et peptidique est vraiment illimité, ce qui fournit un grand nombre de types de protéines dans les cellules vivantes.
Dans les cellules vivantes, les molécules de protéines ou leurs sections individuelles ne constituent pas une chaîne allongée, mais sont tordues en spirale, rappelant un ressort étendu (appelée hélice α) ou pliées en une couche pliée (couche β). Structure secondaire résulte de la formation de liaisons hydrogène entre les groupes -CO- et -NH 2 - de deux liaisons peptidiques au sein d'une chaîne polypeptidique (configuration hélicoïdale) ou entre deux chaînes polypeptidiques (couches repliées).
La protéine kératinique a une configuration complètement hélicoïdale. C'est la protéine structurelle des cheveux, de la fourrure, des ongles, des griffes, du bec, des plumes et des cornes. La structure secondaire en spirale est caractéristique, outre la kératine, de protéines fibrillaires (filiformes) telles que la myosine, le fibrinogène et le collagène.
Dans la plupart des protéines, les sections hélicoïdales et non hélicoïdales de la chaîne polypeptidique se replient en une formation sphérique tridimensionnelle - un globule (caractéristique des protéines globulaires). Un globule d'une certaine configuration est structure tertiaireécureuil. La structure tertiaire est stabilisée par des liaisons ioniques, hydrogène, des liaisons disulfure covalentes (qui se forment entre les atomes de soufre qui composent la cystéine), ainsi que par des interactions hydrophobes. Les interactions hydrophobes sont les plus importantes dans l’émergence de la structure tertiaire ; Dans ce cas, la protéine se replie de telle manière que ses chaînes latérales hydrophobes sont cachées à l'intérieur de la molécule, c'est-à-dire qu'elles sont protégées du contact avec l'eau, et que les chaînes latérales hydrophiles, au contraire, sont exposées à l'extérieur.
De nombreuses protéines avec une structure particulièrement complexe sont constituées de plusieurs chaînes polypeptidiques maintenues ensemble dans la molécule en raison d'interactions hydrophobes, ainsi qu'à l'aide de liaisons hydrogène et ioniques - cela se produit structure quaternaire. Cette structure se retrouve, par exemple, dans la protéine globulaire hémoglobine. Sa molécule se compose de quatre sous-unités polypeptidiques distinctes (protomères), situées dans la structure tertiaire, et d'une partie non protéique - l'hème. Ce n'est que dans une telle structure que l'hémoglobine est capable de remplir sa fonction de transport.
Sous l'influence de divers facteurs chimiques et physiques (traitement à l'alcool, à l'acétone, aux acides, aux alcalis, haute température, irradiation, haute pression, etc.), la structure tertiaire et quaternaire de la protéine change en raison de la rupture des liaisons hydrogène et ioniques. . Le processus de perturbation de la structure native (naturelle) d'une protéine est appelé dénaturation. Dans ce cas, il y a une diminution de la solubilité des protéines, un changement dans la forme et la taille des molécules, une perte d'activité enzymatique, etc. Le processus de dénaturation est parfois réversible, c'est-à-dire que le retour des conditions environnementales normales peut s'accompagner de la restauration spontanée de la structure naturelle de la protéine. Ce processus est appelé renaturation. Il s'ensuit que toutes les caractéristiques de la structure et du fonctionnement d'une macromolécule protéique sont déterminées par sa structure primaire.
En fonction de leur composition chimique, les protéines sont divisées en protéines simples et complexes. À simple inclure des protéines constituées uniquement d’acides aminés, et complexe- contenant une partie protéique et une partie non protéique (prostatique) - des ions métalliques, des glucides, des lipides, etc. Les protéines simples sont l'albumine sérique, les immunoglobulines (anticorps), la fibrine, certaines enzymes (trypsine), etc. Les protéines complexes sont toutes des protéolipides et les glycoprotéines, l'hémoglobine, la plupart des enzymes, etc.
Fonctions des protéines
De construction. Les protéines font partie des membranes cellulaires et des organites cellulaires. Les parois des vaisseaux sanguins, du cartilage, des tendons, des cheveux, des ongles et des griffes des animaux supérieurs sont principalement constituées de protéines.
Catalytique (enzymatique). Les protéines enzymatiques catalysent toutes les réactions chimiques dans le corps. Ils assurent la dégradation des nutriments dans le tube digestif, la fixation du carbone lors de la photosynthèse, les réactions de synthèse matricielle, etc.
Transport. Les protéines sont capables de fixer et de transporter diverses substances. Les albumines sanguines transportent les acides gras, les globulines transportent les ions métalliques et les hormones. L'hémoglobine transporte l'oxygène et le dioxyde de carbone.
Les molécules protéiques qui composent la membrane plasmique participent au transport des substances vers et hors de la cellule.
Protecteur. Elle est réalisée par les immunoglobulines (anticorps) présentes dans le sang, qui assurent la défense immunitaire de l’organisme. Le fibrinogène et la thrombine participent à la coagulation du sang et préviennent les saignements.
Contractif. Elle est assurée par le mouvement des filaments de protéines d'actine et de myosine les uns par rapport aux autres dans les muscles et à l'intérieur des cellules. Le glissement des microtubules, construits à partir de la protéine tubuline, explique le mouvement des cils et des flagelles.
Réglementaire. De nombreuses hormones sont des oligopeptides ou des protéines, par exemple : l'insuline, le glucagon, l'hormone adénocorticotrope, etc.
Récepteur. Certaines protéines intégrées dans la membrane cellulaire sont capables de modifier leur structure en réponse à l'environnement externe. C'est ainsi que les signaux sont reçus de l'environnement extérieur et que les informations sont transmises dans la cellule. Un exemple serait phytochrome- une protéine sensible à la lumière qui régule la réponse photopériodique des plantes, et opsine- composant rhodopsine, un pigment présent dans les cellules de la rétine.
Dans une molécule protéique, les résidus d'acides aminés sont reliés par une liaison dite peptidique. La séquence complète des résidus d'acides aminés dans une telle chaîne est appelée structure primaire de la protéine. Le nombre de résidus dans différentes protéines peut varier de quelques à plusieurs milliers. Petites molécules avec une mol. ceux pesant moins de 10 000 daltons sont appelés peptides, et les plus gros sont appelés protéines. Les protéines contiennent généralement des acides aminés acides et alcalins, de sorte que la molécule protéique a des charges positives et négatives. La valeur du pH à laquelle le nombre de charges négatives est égal au nombre de charges positives est appelée point isoélectrique de la protéine.
Généralement, la chaîne protéique se replie en structures plus complexes. L'oxygène du groupe C=O peut former une liaison hydrogène avec l'hydrogène du groupe N-H situé dans un autre acide aminé. Grâce à ces liaisons hydrogène, la structure secondaire de la protéine est formée. L'un des types de structure secondaire est l'hélice b. Dans celui-ci, chaque oxygène du groupe C=O est lié à l'hydrogène du 4ème groupe NH le long de l'hélice. Il y a 3,6 résidus d'acides aminés par tour d'hélice ; le pas d'hélice est de 0,54 nm.
De nombreuses protéines contiennent ce qu'on appelle. structure b, ou couche b, dans laquelle les chaînes polypeptidiques sont presque complètement dépliées, leurs sections individuelles avec leurs groupes -CO- et -NH- forment des liaisons hydrogène avec d'autres sections de la même chaîne ou une chaîne polypeptidique adjacente.
b-La protéine kératine, qui compose les cheveux et la laine, a une structure en spirale. Lorsqu'ils sont chauffés, les cheveux et la laine mouillés s'étirent facilement, puis reviennent spontanément à leur état d'origine : lorsqu'ils sont étirés, les liaisons hydrogène de l'hélice b sont rompues puis progressivement restaurées.
c-La structure est caractéristique de la fibroïne, principale protéine de soie sécrétée par les chenilles du ver à soie. Contrairement à la laine, la soie est presque inextensible - la structure β est formée de chaînes polypeptidiques allongées et il est presque impossible de l'étirer davantage sans rompre les liaisons covalentes.
Le repliement des protéines ne se limite généralement pas à la structure secondaire. Les résidus d’acides aminés hydrophobes « cherchent » à se cacher de l’environnement aqueux à l’intérieur de la molécule protéique. Une interaction électrostatique est possible entre les groupes latéraux des acides aminés acides et alcalins, qui sont chargés respectivement négativement et positivement. De nombreux résidus d’acides aminés peuvent former des liaisons hydrogène entre eux. Enfin, les résidus d'acides aminés cystéine contenant des groupes SH sont capables de former des liaisons covalentes -S-S- entre eux.
Grâce à toutes ces interactions – hydrophobe, ionique, hydrogène et disulfure – la chaîne protéique forme une configuration spatiale complexe appelée structure tertiaire.
Dans la composition du globule de nombreuses protéines, on peut distinguer des régions compactes individuelles d'une taille d'environ 10 000 à 20 000 daltons. Ils sont appelés domaines. Les sections de la chaîne polypeptidique entre les domaines sont très flexibles, de sorte que la structure entière peut être représentée comme des billes de domaines relativement rigides reliées par des sections intermédiaires flexibles de la structure primaire.
De nombreuses protéines (appelées oligomères) sont constituées non pas d'une, mais de plusieurs chaînes polypeptidiques. Leur combinaison forme la structure quaternaire de la protéine, avec les chaînes individuelles appelées sous-unités. La structure quaternaire est tenue par les mêmes liens que la structure tertiaire. La configuration spatiale d’une protéine (c’est-à-dire sa structure tertiaire et quaternaire) est appelée conformation.
Riz. 4.
La principale méthode permettant d'établir la structure spatiale des protéines et d'autres polymères biologiques est l'analyse par diffraction des rayons X. Récemment, de grands progrès ont été réalisés dans la modélisation informatique des conformations des protéines.
Les liaisons hydrogène, électrostatiques et hydrophobes qui créent les structures secondaires, tertiaires et quaternaires d'une protéine sont moins fortes que la liaison peptidique qui forme la structure primaire. Lorsqu'elles sont chauffées, elles sont facilement détruites et, bien que la protéine conserve intacte sa structure primaire, elle ne peut pas remplir ses fonctions biologiques et devient inactive. Le processus de destruction de la conformation naturelle d'une protéine, accompagné d'une perte d'activité, est appelé dénaturation. La dénaturation est causée non seulement par la chaleur, mais également par des produits chimiques qui perturbent les liaisons des structures secondaires et tertiaires - par exemple l'urée, qui, à des concentrations élevées, détruit les liaisons hydrogène dans le globule protéique.
Les liaisons disulfure -S-S forment des « clips » solides qui réticulent différentes sections d'une chaîne polypeptidique ou de différentes chaînes. Ces liaisons existent, par exemple, dans les kératines, et différentes kératines contiennent des quantités différentes de ces liaisons croisées : cheveux et laine - un peu, cornes, sabots de mammifères et carapaces de tortues - bien plus.
La structure secondaire, tertiaire et quaternaire d'une protéine est déterminée par sa structure primaire. En fonction de la séquence d'acides aminés dans la chaîne polypeptidique, des sections b-hélicoïdales ou b-structurelles se formeront, qui s'intégreront alors spontanément dans une certaine structure tertiaire, et pour certaines protéines, des chaînes individuelles se combineront également pour former un structure quaternaire.
Si vous modifiez la structure primaire d’une protéine, toute sa conformation peut changer de manière significative. Il existe une maladie héréditaire grave - la drépanocytose, dans laquelle l'hémoglobine devient légèrement soluble dans l'eau et les globules rouges acquièrent la forme d'une faucille. La cause de la maladie est le remplacement d'un seul acide aminé sur les 574 qui composent l'hémoglobine humaine (l'acide glutamique, situé à la 6ème place de l'extrémité N-terminale de l'une des chaînes d'hémoglobine chez les personnes normales, est remplacé par la valine dans patients).
Le processus d'association spontanée de sous-unités protéiques en complexes complexes à structure quaternaire est appelé auto-assemblage. La plupart des complexes protéiques à structure quaternaire sont formés précisément par auto-assemblage.
Dans les années 1980, on a découvert que toutes les protéines et complexes protéiques ne se forment pas par auto-assemblage. Il s'est avéré que pour la formation de structures telles que les nucléosomes (complexes de protéines histones avec ADN), les pili bactériens, ainsi que certains complexes enzymatiques complexes, des protéines auxiliaires spéciales appelées chaperons sont utilisées. Les chaperons ne font pas partie de la structure résultante, mais contribuent seulement à sa formation.
Les chaperons servent non seulement à organiser des complexes complexes, mais aident dans certains cas à replier correctement une chaîne polypeptidique. Ainsi, lorsqu'il est exposé à des températures élevées, le nombre de cellules augmente fortement. protéines de choc thermique. Ils se lient aux protéines cellulaires partiellement dénaturées et restaurent leur conformation naturelle.
Pendant longtemps, on a cru qu'une protéine ne pouvait avoir qu'une seule conformation stable dans des conditions données, mais récemment, ce postulat a dû être révisé. La raison de cette refonte était la découverte d'agents pathogènes de ce qu'on appelle. infections neurologiques lentes. Ces infections surviennent chez différentes espèces de mammifères. Il s'agit notamment de la maladie du mouton « scrapie », de la maladie humaine « kuru » (« mort par le rire ») et de la récemment sensationnelle « rage des vaches ». Ils ont beaucoup en commun.
Ils se caractérisent par de graves lésions du système nerveux central. Ainsi, les personnes atteintes de kuru, aux premiers stades de la maladie, éprouvent une instabilité émotionnelle (la majorité rit souvent et sans raison, mais certaines sont dans un état de dépression ou d'agressivité non motivée) et de légers troubles de la coordination des mouvements. Aux stades ultérieurs, les patients ne sont plus capables non seulement de bouger, mais même de s'asseoir sans soutien ou de manger.
L'infection se produit généralement par la nourriture (parfois par le sang). La maladie s’est développée chez les animaux après leur avoir donné de la farine d’os, fabriquée à partir des os d’individus malades. Le Kuru est une maladie des cannibales papous, transmise en mangeant le cerveau de parents décédés (se manger les uns les autres dans ce cas est plus une branche du culte que la cuisine ; cela a une signification rituelle importante).
Toutes ces maladies ont une très longue période d’incubation et se développent lentement. Dans le cerveau des patients, on note le dépôt d'un conglomérat protéique insoluble. Les filaments protéiques insolubles se trouvent dans les vésicules situées à l’intérieur des neurones, ainsi que dans la substance extracellulaire. Il y a une destruction des neurones dans certaines parties du cerveau, notamment dans le cervelet.
Pendant longtemps, la nature des agents responsables de ces maladies est restée mystérieuse et ce n'est qu'au début des années 80 qu'il a été établi que ces agents pathogènes étaient des protéines spéciales d'un poids moléculaire d'environ 30 000 daltons. De tels objets jusqu'ici inconnus de la science sont appelés prions.
Il a été constaté que la protéine prion est codée dans l’ADN de l’organisme hôte. La protéine d'un corps sain contient la même séquence d'acides aminés que la protéine d'une particule de prion infectieuse, mais ne provoque aucun symptôme pathologique. La fonction de la protéine prion est encore inconnue. Les souris chez lesquelles les ingénieurs généticiens ont artificiellement désactivé le gène de cette protéine se sont développées tout à fait normalement, même si elles présentaient quelques anomalies dans le fonctionnement du système nerveux central (pire apprentissage, troubles du sommeil). Dans un corps sain, cette protéine se trouve à la surface des cellules de nombreux organes, notamment dans le cerveau.
Il s’est avéré que la protéine prion dans la particule infectieuse a une conformation différente de celle des cellules normales. Il contient des régions bêta-structurelles, est très résistant à la digestion par les enzymes digestives et a la capacité de former des agrégats insolubles (apparemment, le dépôt de tels agrégats dans le cerveau est à l'origine du développement de la neuropathologie).
Le plus intéressant est que la conformation « normale » de cette protéine se transforme en conformation « pathogène » si la cellule entre en contact avec la protéine « pathogène ». Il s’avère que la protéine « pathogène » « sculpte » à elle seule la structure spatiale de la protéine « normale ». Il dirige son repliement comme une matrice, provoquant l’apparition de plus en plus de molécules de conformation « pathogène » et, à terme, la mort de l’organisme.
On ne sait toujours pas exactement comment cela se produit. Si vous mélangez les formes normales et infectieuses de protéine prion dans un tube à essai, aucune nouvelle molécule infectieuse ne se formera. Apparemment, dans une cellule vivante, il existe des sortes de molécules auxiliaires (probablement des chaperons) qui permettent à la protéine prion de faire son sale boulot.
Le dépôt de conglomérats protéiques insolubles peut également provoquer d’autres maladies nerveuses incurables. La maladie d'Alzheimer n'est pas contagieuse : elle survient à un âge avancé chez les personnes ayant une prédisposition héréditaire. Les patients souffrent d'une détérioration de la mémoire, d'un affaiblissement de l'intelligence, de la démence et, finalement, d'une perte totale des fonctions mentales. La raison du développement de la maladie est le dépôt de ce qu'on appelle dans le cerveau. plaques amyloïdes. Ils sont constitués d'une protéine insoluble - l'amyloïde-β. Il s’agit d’un fragment de protéine précurseur amyloïde, une protéine normale présente chez toutes les personnes en bonne santé. Chez les patients, il est décomposé pour former un peptide amyloïde insoluble.
Des mutations dans divers gènes provoquent le développement de la maladie d'Alzheimer. Naturellement, elle est causée par des mutations dans le gène de la protéine précurseur amyloïde - le précurseur modifié, après clivage, forme de la β-amyloïde insoluble, qui forme des plaques et détruit les cellules cérébrales. Mais la maladie survient également lorsqu’il y a une mutation dans les gènes des protéines qui régulent l’activité des protéases qui coupent la protéine précurseur amyloïde. On ne sait pas exactement comment la maladie se développe dans ce cas : peut-être que la protéine précurseur normale est coupée au mauvais endroit, ce qui conduit à la précipitation du peptide résultant.
La maladie d'Alzheimer se développe très tôt chez les patients atteints du syndrome de Down : ils n'ont pas deux copies du 21e chromosome, comme tout le monde, mais trois. Les patients atteints du syndrome de Down ont une apparence et une démence caractéristiques. Le fait est que le gène de la protéine précurseur amyloïde est situé sur le chromosome 21, une augmentation de la quantité du gène entraîne une augmentation de la quantité de protéine et un excès de protéine précurseur conduit à l'accumulation de β-amyloïde insoluble.
Les protéines se combinent souvent avec d'autres molécules. Ainsi, l'hémoglobine, qui transporte l'oxygène dans le système circulatoire, est constituée d'une partie protéique - la globine, et d'une partie non protéique - l'hème. L'ion Fe2+ fait partie de l'hème. La globine est constituée de quatre chaînes polypeptidiques. En raison de la présence d'hème avec le fer, l'hémoglobine catalyse l'oxydation de diverses substances organiques avec du peroxyde d'hydrogène - par exemple la benzidine. Auparavant, cette réaction, appelée test à la benzidine, était utilisée en médecine légale pour détecter des traces de sang.
Certaines protéines sont chimiquement liées aux glucides et sont appelées glycoprotéines. De nombreuses protéines sécrétées par une cellule animale sont des glycoprotéines - par exemple la transferrine et les immunoglobulines, connues dans les sections précédentes. Cependant, la gélatine, bien qu'elle soit un produit de l'hydrolyse de la protéine de collagène sécrétée, ne contient pratiquement aucun glucide attaché. À l’intérieur de la cellule, les glycoprotéines sont beaucoup moins courantes.
Dans la pratique en laboratoire, de nombreuses méthodes sont utilisées pour déterminer la concentration en protéines. Le plus simple d'entre eux utilise un réactif biuret - une solution alcaline de sel de cuivre divalent. Dans un environnement alcalin, certaines des liaisons peptidiques de la molécule protéique se transforment en forme énol, qui forme des complexes de couleur rouge avec le cuivre divalent. Une autre réaction protéique courante est la coloration de Bradford. Au cours de la réaction, les molécules d'un colorant spécial se lient au globule protéique, ce qui provoque un changement brusque de couleur - la solution passe du brun pâle au bleu vif. Ce colorant - "Coomassie Bright Blue" - était auparavant utilisé pour teindre la laine (et la laine, comme nous le savons, est constituée de protéine de kératine). Enfin, pour déterminer la concentration d'une protéine, vous pouvez utiliser sa capacité à absorber la lumière ultraviolette d'une longueur d'onde de 280 nm (les acides aminés aromatiques phénylalanine, tyrosine et tryptophane l'absorbent). Plus la solution absorbe ce rayonnement ultraviolet, plus elle contient de protéines.
Pour décrire la structure d'une molécule protéique, les concepts de structures primaires, secondaires, tertiaires et quaternaires d'une molécule protéique ont été introduits. Ces dernières années, des concepts tels que la structure supersecondaire sont apparus, qui caractérisent les agrégats énergétiquement préférés de la structure secondaire, et les domaines - les parties d'un globule protéique, qui sont des régions globulaires plutôt isolées.
Le nombre et la séquence des acides aminés ainsi que l'emplacement des liaisons disulfure dans la chaîne polypeptidique déterminent la structure primaire de la protéine. Il existe une relation très étroite entre la structure primaire d’une protéine et sa fonction dans un organisme donné. Pour qu’une protéine remplisse sa fonction inhérente, une séquence très spécifique d’acides aminés est requise dans la chaîne polypeptidique de cette protéine. Même de petits changements dans la structure primaire peuvent modifier considérablement les propriétés d'une protéine et, par conséquent, ses fonctions. Par exemple, les globules rouges des personnes en bonne santé contiennent une protéine appelée hémoglobine avec une certaine séquence d'acides aminés. Une petite proportion de personnes présentent une anomalie congénitale dans la structure de l'hémoglobine : leurs globules rouges contiennent de l'hémoglobine, qui contient dans une position l'acide aminé valine (hydrophobe, non polaire) au lieu de l'acide glutamique (chargé, polaire). Une telle hémoglobine diffère considérablement de la normale par ses propriétés physicochimiques et biologiques. L'apparition d'un acide aminé hydrophobe entraîne l'apparition d'un contact hydrophobe « collant » (les globules rouges se déplacent mal dans les vaisseaux sanguins), à une modification de la forme du globule rouge (de biconcave à en forme de croissant) , ainsi qu'à une détérioration du transfert d'oxygène, etc. Les enfants nés avec cette anomalie meurent dans la petite enfance de la drépanocytose.
Des preuves complètes en faveur de l'affirmation selon laquelle l'activité biologique est déterminée par la séquence d'acides aminés ont été obtenues après la synthèse artificielle de l'enzyme ribonucléase (Merrifield). Un polypeptide synthétisé avec la même séquence d'acides aminés que l'enzyme naturelle avait la même activité enzymatique.
Les recherches menées au cours des dernières décennies ont montré que la structure primaire est génétiquement fixée et détermine à son tour les structures secondaires, tertiaires et quaternaires de la molécule protéique ainsi que sa conformation générale. La première protéine dont la structure primaire a été établie était l'hormone protéique insuline (contient 51 acides aminés). Cela a été réalisé en 1953 par Frederick Sanger. À ce jour, la structure primaire de plus de dix mille protéines a été déchiffrée, mais il s'agit d'un très petit nombre si l'on considère qu'il existe environ 10 12 protéines dans la nature.
Connaissant la structure primaire d'une protéine, vous pouvez écrire avec précision sa formule développée si la protéine est représentée par une chaîne polypeptidique. Si la protéine contient plusieurs chaînes polypeptidiques, elles sont d'abord séparées à l'aide de réactifs spéciaux. Pour déterminer la structure primaire d'une chaîne polypeptidique individuelle, sa composition en acides aminés est déterminée par des méthodes d'hydrolyse utilisant des analyseurs d'acides aminés. Ensuite, à l’aide de méthodes et de réactifs spéciaux, la nature des acides aminés terminaux est déterminée. Pour établir l'ordre d'alternance des acides aminés, la chaîne polypeptidique est soumise à une hydrolyse enzymatique, qui produit des fragments de cette chaîne polypeptidique - des peptides courts. Ces peptides sont séparés par chromatographie et la séquence d'acides aminés de chacun est déterminée. Ainsi, on atteint un stade où la séquence des acides aminés dans les peptides individuels (fragments protéiques) est connue, mais la séquence des peptides eux-mêmes reste floue. Cette dernière est établie à l’aide de peptides dits chevauchants. Pour ce faire, une autre enzyme est utilisée, qui clive la chaîne polypeptidique d'origine dans d'autres sections, et la séquence d'acides aminés des peptides nouvellement obtenus est déterminée. Les peptides formés sous l'action de deux enzymes contiennent des fragments identiques de séquences d'acides aminés. En les combinant, la séquence générale d'acides aminés de la chaîne polypeptidique est établie.
L. Pauling et R. Corey ont apporté une grande contribution à l'étude de la structure de la molécule protéique. Remarquant que la molécule protéique contient le plus grand nombre de liaisons peptidiques, ils ont été les premiers à mener des études minutieuses aux rayons X de cette liaison. Nous avons étudié les longueurs des liaisons, les angles selon lesquels les atomes sont situés et la direction des atomes par rapport à la liaison. Sur la base de la recherche, les principales caractéristiques suivantes de la liaison peptidique ont été établies.
1. Les quatre atomes de la liaison peptidique et les deux atomes de carbone α qui y sont attachés se trouvent dans le même plan. Les groupes d’atomes de carbone R et H se trouvent en dehors de ce plan.
2. Les atomes O et H de la liaison peptidique et les deux atomes de carbone α et les groupes R ont une orientation trans par rapport à la liaison peptidique.
3. La longueur de la liaison C-N, égale à 1,32 Å, est intermédiaire entre la longueur d'une double liaison covalente (1,21 Å) et d'une liaison covalente homogène (1,47 Å). Il s'ensuit que la liaison C-N présente en partie le caractère d'une double liaison. Ceux. la liaison peptidique peut exister sous forme de structures de résonance et tautamériques, sous forme céto-énol.

La rotation autour de la liaison –C=N– est difficile et tous les atomes inclus dans le groupe peptidique ont une configuration trans planaire. La configuration cis est énergétiquement moins favorable et ne se retrouve que dans certains peptides cycliques. Chaque fragment peptidique plan contient deux liaisons avec des atomes de carbone α capables de rotation. Il s'agit des liaisons C –N (l'angle de rotation autour de cette liaison est noté ) et d'une liaison C –C (l'angle de rotation autour de cette liaison est noté ).
La liaison peptidique est de nature chimique covalente et confère une résistance élevée à la structure primaire de la molécule protéique. Étant un élément répétitif de la chaîne polypeptidique et possédant des caractéristiques structurelles spécifiques, la liaison peptidique affecte non seulement la forme de la structure primaire, mais également les niveaux supérieurs d'organisation de la chaîne polypeptidique.
La structure secondaire d'une molécule protéique est formée à la suite de l'un ou l'autre type de rotation libre autour des liaisons reliant les atomes de carbone α dans la chaîne polypeptidique.
Trois principaux types de structure se trouvent dans les chaînes polypeptidiques naturelles : l’hélice α, la feuille pliée et la bobine aléatoire. Une structure hélicoïdale est formée si la chaîne a les mêmes angles de rotation () pour toutes les liaisons C –N et angles de rotation () pour toutes les liaisons C –C et sont égaux à –48º et –57º, respectivement. On trouve l'hélice α droitière la plus courante. Cette structure est très stable car il a peu ou pas d'encombrement stérique, en particulier pour les groupes R des chaînes latérales d'acides aminés. Les groupes R des acides aminés sont dirigés vers l'extérieur à partir de l'axe central de l'hélice α. Les dipôles de l'hélice β = C = O et N – H des liaisons peptidiques voisines sont orientés de manière optimale (presque coaxiale) pour l'interaction dipolaire, formant ainsi un système étendu de liaisons hydrogène coopératives intramoléculaires qui stabilisent l'hélice α. Le pas de l'hélice (un tour complet) est de 5,4Å et comprend 3,6 résidus d'acides aminés.

Figure 1 – Structure et paramètres de l’hélice α de la protéine
La structure en spirale peut être perturbée par deux facteurs :
1) en présence d'un résidu proline dont la structure cyclique introduit une rupture dans la chaîne peptidique - il n'y a pas de groupe –NH 2, donc la formation d'une liaison hydrogène intrachaîne est impossible ;

2) si dans une chaîne polypeptidique il y a de nombreux résidus d'acides aminés consécutifs qui ont une charge positive (lysine, arginine) ou une charge négative (acides glutamique, aspartique), dans ce cas, il y a une forte répulsion mutuelle de groupes chargés de manière similaire (– COO– ou –NH 3 +) dépasse largement l'influence stabilisatrice des liaisons hydrogène dans l'hélice β.
La structure de type feuille pliée est également stabilisée par des liaisons hydrogène entre les mêmes dipôles =NH...... O=C. Cependant, dans ce cas, une structure complètement différente apparaît, dans laquelle le squelette de la chaîne polypeptidique est allongé de telle manière qu'il présente une structure en zigzag. Les angles de rotation des liaisons С -N () et С -С () sont proches de –120+135 0, respectivement. Les régions repliées de la chaîne polypeptidique présentent des propriétés coopératives, c'est-à-dire ont tendance à être situés côte à côte dans une molécule protéique et à former des

des chaînes polypeptidiques identiques ou antiparallèles,

qui sont renforcés grâce aux liaisons hydrogène entre ces chaînes. De telles structures sont appelées feuilles pliées en (Figure 2).

Figure 2 - Structure des chaînes polypeptidiques
-Les feuilles pliées en spirale sont des structures ordonnées ; elles ont une disposition régulière de résidus d'acides aminés dans l'espace. Les sections d'une chaîne protéique avec une disposition irrégulière de résidus d'acides aminés dans l'espace, qui sont également maintenues ensemble grâce aux liaisons hydrogène, sont appelées désordonnées, sans structure - une bobine statistique. Toutes ces structures apparaissent spontanément et automatiquement du fait qu'un polypeptide donné possède une certaine séquence d'acides aminés, génétiquement prédéterminée. Les hélices et les structures déterminent une certaine capacité des protéines à remplir des fonctions biologiques spécifiques. Ainsi, la structure α-hélicoïdale (α-kératine) est bien adaptée pour former des structures protectrices externes - plumes, cheveux, cornes, sabots. La structure α favorise la formation de fils flexibles et inextensibles de soie et de toiles d'araignées, ainsi que la conformation de ceux-ci. la protéine de collagène assure une grande résistance à la rupture nécessaire aux tendons. La présence uniquement d'hélices α ou de structures β est caractéristique des protéines filamenteuses-fibrillaires. Dans la composition des protéines globulaires sphériques, la teneur en hélices α, en structures β et en régions sans structure varie considérablement. Par exemple : l'insuline est spiralée à 60 %, l'enzyme ribonucléase à 57 %, le lysozyme de la protéine d'œuf de poule à 40 %.
Les informations sur l'alternance des résidus d'acides aminés dans une chaîne polypeptidique, ainsi que sur la présence de sections spiralées, pliées et désordonnées dans une molécule protéique, ne fournissent pas encore une image complète ni du volume ni de la forme, et encore moins de la position relative de sections de la chaîne polypeptidique les unes par rapport aux autres.
Ces caractéristiques structurelles d'une protéine sont clarifiées en étudiant sa structure tertiaire, comprise comme la disposition spatiale générale dans un certain volume de la chaîne polypeptidique.
La structure tertiaire est établie par analyse par diffraction des rayons X. Le premier modèle d'une molécule protéique, la myoglobine, reflétant sa structure tertiaire, a été créé par J. Kendrew et ses collaborateurs en 1957. Malgré de grandes difficultés, il a été possible à ce jour d'établir la structure tertiaire de plus de 1000 protéines, dont l'hémoglobine, la pepsine, le lysozyme, l'insuline, etc.
La structure tertiaire des protéines est formée par repliement supplémentaire de la chaîne peptidique contenant une hélice α, une structure β et des régions sans structure périodique. La structure tertiaire d'une protéine se forme de manière entièrement automatique, spontanée et est entièrement prédéterminée par la structure primaire. Le principal moteur de l’émergence d’une structure tridimensionnelle est l’interaction des radicaux d’acides aminés avec les molécules d’eau. Dans ce cas, les radicaux d’acides aminés hydrophobes non polaires sont regroupés à l’intérieur de la molécule protéique, tandis que les radicaux polaires sont orientés vers l’eau. À un moment donné, la conformation stable thermodynamiquement la plus favorable de la molécule, un globule, apparaît. Sous cette forme, la molécule protéique se caractérise par une énergie libre minimale. La conformation du globule résultant est influencée par des facteurs tels que le pH de la solution, la force ionique de la solution, ainsi que l'interaction des molécules protéiques avec d'autres substances.
Récemment, des preuves sont apparues selon lesquelles le processus de formation de structures tertiaires n’est pas automatique, mais est régulé et contrôlé par des mécanismes moléculaires spéciaux. Ce processus implique des protéines spécifiques – les chaperons. Leurs fonctions principales sont la capacité d'empêcher la formation de bobines aléatoires non spécifiques (chaotiques) à partir de la chaîne polypeptidique et d'assurer leur livraison (transport) vers des cibles subcellulaires, créant ainsi les conditions nécessaires à l'achèvement du repliement de la molécule protéique.
La stabilisation de la structure tertiaire est assurée grâce à des interactions non covalentes entre groupes atomiques de radicaux latéraux des types suivants :
des liaisons hydrogène peuvent se produire entre des groupes fonctionnels de radicaux latéraux. Par exemple, entre le groupe OH de la tyrosine et –N dans le cycle d'un résidu histidine.
forces d'attraction électrostatiques entre des radicaux portant des groupes ioniques de charges opposées (interactions ion-ion), par exemple, le groupe carboxyle chargé négativement (– COO –) de l'acide aspartique et le groupe α-amino chargé positivement (NH 3 +) d'une lysine résidu.
Les interactions hydrophobes sont provoquées par les forces de Van der Waals entre les radicaux d'acides aminés non polaires. (Par exemple, les groupes –CH 3 – alanine.
La structure tertiaire est également stabilisée par des liaisons disulfure covalentes (–S–S–) entre les résidus de cystéine. Cette liaison est très forte et n’est pas présente dans toutes les protéines. Cette connexion joue un rôle important dans les substances protéiques des céréales et de la farine, car influence la qualité du gluten, les propriétés structurelles et mécaniques de la pâte et, par conséquent, la qualité du produit fini - pain, etc.
Un globule protéique n'est pas une structure absolument rigide : dans certaines limites, des mouvements réversibles de parties de la chaîne peptidique les unes par rapport aux autres sont possibles avec la rupture d'un petit nombre de liaisons faibles et la formation de nouvelles. La molécule semble respirer, pulser dans ses différentes parties. Ces pulsations ne perturbent pas le plan de conformation de base de la molécule, tout comme les vibrations thermiques des atomes d'un cristal ne modifient pas la structure du cristal si la température n'est pas si élevée qu'elle fond.
Ce n’est qu’après qu’une molécule protéique a acquis une structure tertiaire naturelle et native qu’elle présente son activité fonctionnelle spécifique : catalytique, hormonale, antigénique, etc. C'est lors de la formation de la structure tertiaire que se produit la formation de centres actifs d'enzymes, centres responsables de l'intégration des protéines dans le complexe multienzymatique, centres responsables de l'auto-assemblage des structures supramoléculaires. Ainsi, tout effet (thermique, physique, mécanique, chimique) conduisant à la destruction de cette conformation native de la protéine (rupture des liaisons) s’accompagne d’une perte partielle ou totale des propriétés biologiques de la protéine.
L'étude des structures chimiques complètes de certaines protéines a montré que dans leur structure tertiaire sont identifiées des zones où se concentrent les radicaux d'acides aminés hydrophobes, et que la chaîne polypeptidique est en fait enroulée autour du noyau hydrophobe. De plus, dans certains cas, deux, voire trois noyaux hydrophobes sont séparés dans une molécule protéique, ce qui donne une structure à 2 ou 3 noyaux. Ce type de structure moléculaire est caractéristique de nombreuses protéines ayant une fonction catalytique (ribonucléase, lysozyme…). Une partie ou une région distincte d’une molécule protéique qui possède un certain degré d’autonomie structurelle et fonctionnelle est appelée un domaine. Un certain nombre d'enzymes, par exemple, ont des domaines distincts de liaison au substrat et de liaison aux coenzymes.
La structure tertiaire d’une protéine est directement liée à sa forme, qui peut varier de sphérique à filamenteuse. La forme d'une molécule de protéine est caractérisée par un indicateur tel que le degré d'asymétrie (le rapport entre l'axe long et l'axe court). Les protéines fibrillaires ou filamenteuses ont un degré d'asymétrie supérieur à 80. Avec un degré d'asymétrie inférieur à 80, les protéines sont classées comme globulaires. La plupart d'entre eux ont un degré d'asymétrie de 3 à 5, c'est-à-dire la structure tertiaire est caractérisée par un emballage assez dense de la chaîne polypeptidique, se rapprochant de la forme d'une boule.
Biologiquement, les protéines fibrillaires jouent un rôle très important lié à l’anatomie et à la physiologie des animaux. Chez les vertébrés, ces protéines représentent 1/3 de leur contenu total. Un exemple de protéines fibrillaires est la fibroïne, protéine de soie, qui se compose de plusieurs chaînes antiparallèles avec une structure en feuille pliée. La protéine α-kératine contient 3 à 7 chaînes. Le collagène a une structure complexe dans laquelle 3 chaînes lévogyres identiques sont tordues ensemble pour former une triple hélice dextrogyre. Cette triple hélice est stabilisée par de nombreuses liaisons hydrogène intermoléculaires. La présence d'acides aminés tels que l'hydroxyproline et l'hydroxylysine contribue également à la formation de liaisons hydrogène qui stabilisent la structure de la triple hélice. Toutes les protéines fibrillaires sont peu solubles ou complètement insolubles dans l'eau, car elles contiennent de nombreux acides aminés contenant des groupes R hydrophobes et insolubles dans l'eau : isoleucine, phénylalanine, valine, alanine et méthionine. Après un traitement spécial, le collagène insoluble et indigeste est transformé en un mélange polypeptidique soluble dans la gélatine, qui est ensuite utilisé dans l'industrie alimentaire.
Les protéines globulaires remplissent diverses fonctions biologiques. Ils remplissent une fonction de transport, c'est-à-dire transporter des nutriments, des ions inorganiques, des lipides, etc. Les hormones, ainsi que les composants des membranes et des ribosomes, appartiennent à la même classe de protéines. Toutes les enzymes sont également des protéines globulaires.
Les protéines contenant deux ou plusieurs chaînes polypeptidiques sont appelées protéines oligomères ; elles se caractérisent par la présence d'une structure quaternaire. Les chaînes polypeptidiques (dimensions) de ces protéines peuvent être identiques ou différentes. Les protéines oligomères sont dites homogènes si leurs protomères sont identiques et hétérogènes si leurs protomères sont différents. Par exemple, la protéine hémoglobine est constituée de 4 chaînes : deux protomères - et deux -. L'enzyme α-amylase est constituée de 2 chaînes polypeptidiques identiques. Dans les protéines oligomères, chacune des chaînes polypeptidiques est caractérisée par sa structure secondaire et tertiaire et est appelée sous-unité ou protomère. Les protomères interagissent entre eux non pas avec n'importe quelle partie de leur surface, mais avec une certaine zone (surface de contact). Les surfaces de contact présentent un tel agencement de groupes atomiques entre lesquels naissent des liaisons hydrogène, ioniques et hydrophobes. De plus, la géométrie des protomères favorise également leur connexion. Les protomères s’emboîtent comme la clé d’une serrure. De telles surfaces sont dites complémentaires. Chaque protomère interagit avec l’autre en plusieurs points, rendant impossible la connexion avec d’autres chaînes polypeptidiques ou protéines. De telles interactions complémentaires de molécules sont à la base de tous les processus biochimiques du corps. La structure quaternaire fait référence à la disposition des chaînes polypeptidiques (protomères) les unes par rapport aux autres, c'est-à-dire un procédé de leur repliement et de leur conditionnement conjoints pour former la conformation native d'une protéine oligomère, à la suite de laquelle la protéine a l'une ou l'autre activité biologique.